Yu Huang, Siqi Hu, Fei Guo,NHC Key Laboratory of Systems Biology of Pathogens, Institute of Pathogen Biology, Chinese Academy of Medical Sciences & Peking Union Medical College, Beijing 100730, China
Abstract Stress granule (SG) formation is a primary mechanism through which gene expression is rapidly modulated when the eukaryotic cells undergo cellular stresses (including heat shock, oxidative stress, starvation, viral infection). SGs have been proposed to affect mRNA translation and stability, as well as being linked to apoptosis and nuclear processes. Formation of SGs after viral infection result in blockade of viral protein synthesis and viral replication. Not surprisingly, viruses from diverse families have been found to modulate SG formation in infected cells by associating with important SG effector proteins. Here we provide a summary of the current understanding of the mechanism of SG formation, describe the current knowledge on viruses induce and/or modulate SGs in infected cells via phosphorylation of eIF2α, and regulation of SGs in virus systems. Further, we summarize recent progresses in understanding the relationship between viruses and stress granules in mammalian cells, and suggest that SG formation is an important aspect of the antiviral innate immune response. Keywords:stress granules;translation arrest;virus;innate immunity
PDF (500KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 黄羽, 胡斯奇, 郭斐. 应激颗粒与病毒的相互制约[J]. 遗传, 2019, 41(6): 494-508 doi:10.16288/j.yczz.19-020 Yu Huang, Siqi Hu, Fei Guo. Interaction between stress granules and viruses[J]. Hereditas(Beijing), 2019, 41(6): 494-508 doi:10.16288/j.yczz.19-020
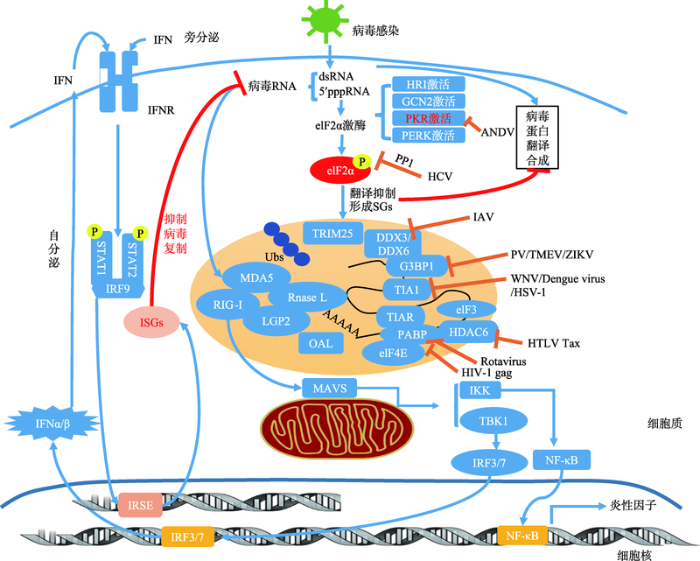
正粘病毒科(Orthomyxoviridae)家族为一类包含反向ssRNA基因组的包膜病毒。流感病毒A (influenza A virus, IAV)通过表达非结构蛋白NS1来抑制PKR的活性,进而抑制SGs的聚集,如感染NS1缺失或突变的IAV,则可诱导形成SGs[86]。NS1介导的SGs的抑制作用,依赖于NS1于RNA相关蛋白55 (RNA associated protein 55, RAP55)的相互作用[87]。除了NS1、IAV的NP和PA-X都能通过非elF2α磷酸化依赖的方式来抵抗SGs的聚集[88]。此外,当感染IAV后,DDX3可与NP相互作用,并调节IFN的产生及SGs的聚集;当感染宿主的IAV缺失NS1,DDX3还能够与产生的SGs共定位[89];DDX6也可结合病毒RNA,促进RIG-I介导的干扰素应答反应,起到抗病毒作用[90]。
目前已知SGs在许多层面上都与先天性免疫相互关联,因此,在对于应激颗粒的研究中可能会发现一些在抗病毒治疗中具有价值的广谱的作用位点。由药物诱发的,经PKR或elF2α磷酸化生成的SGs,有望控制病毒感染。体外实验已经证明,elF4A解旋酶抑制剂hippuristanol可抑制卡里色病毒(caliciviruses)[120];pateamine A可抑制流感病毒A (influenza A virus)的复制[88]。不过,这些药物对于未感染细胞的毒性影响限制了其发展。
BuchanJR, ParkerR . Eukaryotic stress granules: the ins and out of translation Mol Cell, 2009,36(6):932-941. URL [本文引用: 1]
Kedersha NL. GuptaM, LiW, MillerI, AndersonP . RNA-binding proteins TIA-1 and TIAR link the phosphorylation of eIF-2α to the assembly of mammalian stress granules J Cell Biol, 1999,147(7):1431-1442. URL [本文引用: 3]
JacksonRJ, HellenCU, PestovaTV . The mechanism of eukaryotic translation initiation and principles of its regulation Nat Rev Mol Cell Biol, 2010,11(2):113-127. [本文引用: 2]
OkonskiKM, SamuelCE . Stress granule formation induced by measles virus is protein kinase PKR dependent and impaired by RNA adenosine deaminase ADAR1 J Virol, 2012,87(2):756-766. [本文引用: 2]
OnomotoK, JogiM, YooJS, NaritaR, MorimotoS, TakemuraA, SambharaS, KawaguchiA, OsariS, NagataK, MatsumiyaT, NamikiH, YoneyamaM, FujitaT . Critical role of an antiviral stress granule containing RIG-I and PKR in viral detection and innate immunity PLoS One, 2012,7(8):e43031. URL [本文引用: 2]
LuL, HanAP, ChenJJ . Translation initiation control by heme-regulated eukaryotic initiation factor 2alpha kinase in erythroid cells under cytoplasmic stresses Mol Cell Biol, 2001,21(23):7971-7980. URL [本文引用: 1]
Sheree A. WekSZ, WekRC . The histidyl-tRNA synthetase-related sequence in the elf-2α protein kinase GCN2 interacts with tRNA and is required for activation in response to starvation for different amino acids Mol Cell Biol, 1995,15(8):4497-4506. URL [本文引用: 1]
Jing DHPH, BrianR . Activation of GCN2 in UV-irradiated cells inhibits translation Curr Biol, 2002,12(15):1279-1286. URL [本文引用: 1]
GarcíaMA, MeursEF, EstebanM . The dsRNA protein kinase PKR: virus and cell control Biochimie, 2007,89(6-7):799-811. URL [本文引用: 1]
HardingHP, ZhangY, BertolottiA, ZengH, RonD . Perk is essential for translational regulation and cell survival during the unfolded protein response Mol Cell, 2000,5(5):897-904. URL [本文引用: 1]
ZoncuR, EfeyanA, SabatiniDM . MTOR: from growth signal integration to cancer, diabetes and ageing Nat Rev Mol Cell Biol, 2011,12(1):21-35. [本文引用: 1]
von der HaarT, GrossJD, WagnerG, McCarthyJE . The mRNA cap-binding protein eIF4E in post-transcriptional gene expression Nat Struct Mol Biol, 2004,11(6):503-511. [本文引用: 1]
PanniersR . Translational control during heat shock Biochimie, 1994,76:737-747. URL [本文引用: 1]
ThomasMG, LoschiM, DesbatsMA, BoccaccioGL . RNA granules: the good, the bad and the ugly Cell Signal, 2011,23(2):324-334. URL [本文引用: 1]
KwonS, ZhangY, MatthiasP . The deacetylase HDAC6 is a novel critical component of stress granules involved in the stress response Gene Dev, 2007,21(24):3381-3394. URL [本文引用: 1]
LeungAK, VyasS, RoodJE, BhutkarA, SharpPA, ChangP . Poly(ADP-ribose) regulates stress responses and microRNA activity in the cytoplasm Mol Cell, 2011,42(4):489-499. URL [本文引用: 1]
OhnT, KedershaN, HickmanT, TisdaleS, AndersonP . A functional RNAi screen links O-GlcNAc modification of ribosomal proteins to stress granule and processing body assembly Nat Cell Biol, 2008,10(10):1224-1231. [本文引用: 1]
KedershaN, IvanovP, AndersonP . Stress granules and cell signaling: more than just a passing phase? Trends Biochem Sci, 2013,38(10):494-506. URL [本文引用: 1]
BoeynaemsS, AlbertiS, FawziNL, MittagT, PolymenidouM, RousseauF, SchymkowitzJ, ShorterJ, WolozinB, Van Den BoschL, TompaP, FuxreiterM . Protein phase separation: a new phase in cell biology Trends Cell Biol, 2018,28(6):420-435. URL [本文引用: 1]
TakaharaT, MaedaT . Transient sequestration of TORC1 into stress granules during heat stress Mol Cell, 2012,47(2):242-252. URL [本文引用: 1]
BuchanR, KolaitisRM, TaylorJP, ParkerR . Eukaryotic stress granules are cleared by autophagy and Cdc48VCP function Cell, 2013,153(7):1461-1474. URL [本文引用: 1]
ChitiproluM, JagowC, TremblayV, Bondy-ChorneyE, ParisG, SavardA, PalidworG, BarryFA, ZinmanL, KeithJ, RogaevaE, RobertsonJ, Lavallée-AdamM, WoulfeJ, CoutureJF, C?téJ, GibbingsD . A complex of C9ORF72 and p62 uses arginine methylation to eliminate stress granules by autophagy Nat Commun, 2018,9(1):2794. [本文引用: 1]
McInerneyGM, KedershaNL, KaufmanRJ, AndersonP, Liljestr?mP . Importance of eIF2alpha phosphorylation and stress granule assembly in alphavirus translation regulation Mol Biol Cell, 2005,16(8):3753-3763. URL [本文引用: 1]
SciortinoMT, ParisiT, SiracusanoG, MastinoA, TaddeoB, RoizmanB . The virion host shutoff RNase plays a key role in blocking the activation of protein kinase R in cells infected with herpes simplex virus 1 J Virol, 2013,87(6):3271-3276. URL [本文引用: 1]
CassadyKA, GrossM . The Herpes Simplex virus type 1 U(S)11 protein interacts with protein kinase R in infected cells and requires a 30-amino-acid sequence adjacent to a kinase substrate domain J Virol, 2002,76(5):2029-2035. URL [本文引用: 1]
HeB, GrossM, RoizmanB . The γ134.5 protein of herpes simplex virus 1 complexes with protein phosphatase 1α to dephosphorylate the α subunit of the eukaryotic translation initiation factor 2 and preclude the shutoff of protein synthesis by double-stranded RNA- activated protein kinase Proc Natl Acad Sci USA, 1996,94(3):843-848. [本文引用: 1]
MulveyM, AriasC, MohrI . Maintenance of endoplasmic reticulum (ER) homeostasis in herpes simplex virus type 1-infected cells through the association of a viral glycoprotein with PERK, a cellular ER stress sensor J Virol, 2007,81(7):3377-3390. URL [本文引用: 1]
EsclatineA, TaddeoB, RoizmanB . Herpes simplex virus 1 induces cytoplasmic accumulation of TIA-1/TIAR and both synthesis and cytoplasmic accumulation of tristetraprolin, two cellular proteins that bind and destabilize AU-rich RNAs J Virol, 2004,78(16):8582-8592. URL [本文引用: 1]
DauberB, PoonD, Dos SantosT, DuguayBA, MehtaN, SaffranHA, SmileyJR . The herpes simplex virus virion host shutoff protein enhances translation of viral true late mRNAs independently of suppressing protein kinase R and stress granule formation J Virol, 2016,90(13):6049-6057. URL [本文引用: 1]
BurgessHM, MohrI . Defining the role of stress granules in innate immune suppression by the herpes simplex virus 1 endoribonuclease VHS J Virol, 2018,92(15):e00829-18. [本文引用: 1]
KatsafanasGC, MossB . Vaccinia virus intermediate stage transcription is complemented by Ras-GTPase- activating protein SH3 domain-binding protein (G3BP) and cytoplasmic activation/proliferation-associated protein (p137) individually or as a heterodimer J Biol Chem, 2004,279(50):52210-52217. URL [本文引用: 2]
KatsafanasGC, MossB . Linkage of transcription and translation within cytoplasmic poxvirus DNA factories provides a mechanism to coordinate viral and usurp host functions Cell Host Microbe, 2007,2(4):221-228. URL [本文引用: 1]
ZaborowskaI, KellnerK, HenryM, MeleadyP, WalshD . Recruitment of host translation initiation factor eIF4G by the Vaccinia Virus ssDNA-binding protein I3 Virology, 2012,425(1):11-22. URL [本文引用: 1]
MonteroH, RojasM, AriasCF, LópezS . Rotavirus infection induces the phosphorylation of eIF2alpha but prevents the formation of stress granules J Virol, 2008,82(3):1496-1504. URL [本文引用: 1]
RojasM, AriasCF, LópezS . Protein kinase R is responsible for the phosphorylation of eIF2α in rotavirus infection J Virol, 2010,84(20):10457-10466. URL [本文引用: 1]
QinQ, HastingsC, MillerCL . Mammalian orthoreovirus particles induce and are recruited into stress granules at early times postinfection J Virol, 2009,83(21):11090-11101. URL [本文引用: 1]
QinQ, CarrollK, HastingsC, MillerCL . Mammalian orthoreovirus escape from host translational shutoff correlates with stress granule disruption and is independent of eIF2alpha phosphorylation and PKR J Virol, 2011,85(17):8798-8810. URL [本文引用: 1]
CarrollK, HastingsC, MillerCL . Amino acids 78 and 79 of mammalian orthoreovirus protein μNS are necessary for stress granule localization, core proteinλ2 interaction, and de novo virus replication Virology, 2014,448:133-145. URL [本文引用: 1]
FungG, NgCS, ZhangJ, ShiJ, WongJ, PiesikP, HanL, ChuF, JagdeoJ, JanE, FujitaT, LuoH . Production of a dominant-negative fragment due to G3BP1 cleavage contributes to the disruption of mitochondria-associated protective stress granules during CVB3 infection PLoS One, 2013,8(11):e79546. URL [本文引用: 1]
BorgheseF, MichielsT . The leader protein of cardioviruses inhibits stress granule assembly J Virol, 2011,85(18):9614-9622. URL [本文引用: 1]
ReinekeLC, KedershaN. LangereisMA, van KuppeveldFJ, LloydRE , Stress granules regulate double-stranded RNA-dependent protein kinase activation through a complex containing G3BP1 and Caprin1 MBio, 2015,6(2):e02486. [本文引用: 1]
VisserLG, MedinaGNRabouwHH, de GrootRG, LangereisMA, de los SantosT, van KuppeveldFJM . Foot-and-Mouth disease leader protease cleaves G3BP1 and G3BP2 and inhibits stress granule formation J Virol, 2019,93(2):e00922-18. [本文引用: 1]
LiW, LiY, KedershaN, AndersonP, EmaraM, SwiderekKM, MorenoGT, BrintonMA . Cell proteins TIA-1 and TIAR interact with the 3' Stem-Loop of the West nile virus complementary minus-strand RNA and facilitate virus replication J Virol, 2002,76(23):11989-12000. URL [本文引用: 1]
CourtneySC, ScherbikSV, StockmanBM, BrintonMA . West nile virus infections suppress early viral RNA synthesis and avoid inducing the cell stress granule response J Virol, 2012,86(7):3647-3657. URL [本文引用: 1]
ShivesKD, BeatmanEL, ChamanianMO'BrienC, Hobson-PetersJ, BeckhamJD , . West nile virus-induced activation of mammalian target of rapamycin complex 1 supports viral growth and viral protein expression J Virol, 2014,88(16):9458-9471. URL [本文引用: 1]
XiaJ, ChenX, XuF, WangY, ShiY, LiY, HeJ, ZhangP . Dengue virus infection induces formation of G3BP1 granules in human lung epithelial cells Arch Virol, 2015,160(12):2991-2999. URL [本文引用: 1]
BidetK, DadlaniD, Garcia-BlancoMA . G3BP1, G3BP2 and CAPRIN1 are required for translation of interferon stimulated mRNAs and are targeted by a dengue virus non-coding RNA PLoS Pathog, 2014,10(7):e1004242. URL [本文引用: 1]
ZhangHW, MengXY, LiLF, YangYY, QiuHJ . Long non-coding RNAs: Emerging regulators of antiviral innate immune responses Hereditas(Beijing), 2018,40(7):525-533 [本文引用: 1]
MokBWY, SongW, WangP, TaiH, ChenY, ZhengM, WenX, LauSY, WuWL, MatsumotoK, YuenKY, ChenH . The NS1 protein of influenza a virus interacts with cellular processing bodies and stress granules through RNA-associated protein 55 (RAP55) during virus infection J Virol, 2012,86(23):12695-12707. URL [本文引用: 1]
LindquistME, MainouBA, DermodyTS, Crowe JEJr . Activation of protein kinase R is required for induction of stress granules by respiratory syncytial virus but dispensable for viral replication Virology, 2011,413(1):103-110. URL [本文引用: 1]
LiflandAW, JungJ, AlonasE, ZurlaC, Crowe JEJr, SantangeloPJ . Human respiratory syncytial virus nucleoprotein and inclusion bodies antagonize the innate immune response mediated by MDA5 and MAVS J Virol, 2012,86(15):8245-8258. URL [本文引用: 1]
FrickeJ, KooLY, BrownCR, CollinsPL . P38 and OGT sequestration into viral inclusion bodies in cells infected with human respiratory syncytial virus suppresses MK2 activities and stress granule assembly J Virol, 2012,87(3):1333-1347. [本文引用: 1]
RandallRE, GoodbournS . Interferons and viruses: an interplay between induction, signalling, antiviral responses and virus countermeasures J Gen Virol, 2008,89(Pt 1):1-47. URL [本文引用: 1]
IseniF, GarcinD, NishioM, KedershaN, AndersonP, KolakofskyD . Sendai virus trailer RNA binds TIAR, a cellular protein involved in virus-induced apoptosis Embo J, 2002,21(19):5141-5150. URL [本文引用: 1]
Valiente-EcheverríaF, HermosoMA, Soto-RifoR . RNA helicase DDX3: at the crossroad of viral replication and antiviral immunity Rev Med Virol, 2015,25(5):286-299. URL [本文引用: 1]
MirMA, DuranWA, HjelleBL, YeC, PanganibanAT . Storage of cellular 5′ mRNA caps in P bodies for viral cap-snatching Proc Natl Acad Sci USA, 2008,105(49):19294-19299. URL [本文引用: 1]
CimicaV, DalrympleNA, RothE, NasonovA, MackowER . An innate immunity-regulating virulence determinant is uniquely encoded by the Andes virus nucleocapsid protein MBio, 2014,5(1):e01088-13. [本文引用: 1]
MatthysVS, CimicaV, DalrympleNA, GlennonNB, BiancoC, MackowER . Hantavirus GnT elements mediate TRAF3 binding and inhibit RIG-I/TBK1-directed beta interferon transcription by blocking IRF3 phosphorylation J Virol, 2014,88(4):2246-2259. URL [本文引用: 1]
WangZ, MirMA . Andes virus nucleocapsid protein interrupts protein kinase R dimerization to counteract host interference in viral protein synthesis J Virol, 2015,89(3):1628-1639. URL [本文引用: 1]
FangJ, PietzschC, RamanathanP, SantosRI, IlinykhPA, Garcia-BlancoMA, BukreyevA, BradrickSS . Staufen1 interacts with multiple components of the ebola virus ribonucleoprotein and enhances viral RNA synthesis MBio, 2018,9(5):e01771-18. [本文引用: 1]
LegrosS, BoxusM, GatotJS, Van LintC, KruysV, KettmannR, TwizereJC, DequiedtF . The HTLV-1 tax protein inhibits formation of stress granules by interacting with histone deacetylase 6 Oncogene, 2011,30(38):4050-4062. [本文引用: 1]
TakahashiM, HiguchiM, MakokhaGN, MatsukiH, YoshitaM, TanakaY, FujiiM . HTLV-1 tax oncoprotein stimulates ROS production and apoptosis in T cells by interacting with USP10 Blood, 2013,122(5):715-725. URL [本文引用: 1]
Henao-MejiaJ, LiuY, ParkIW, ZhangJ, SanfordJ, HeJJ . Suppression of HIV-1 Nef translation by Sam68 mutant-induced stress granules and nef mRNA sequestration Mol Cell, 2009,33(1):87-96. URL [本文引用: 1]
Valiente-EcheverríaF, MelnychukL, VybohK, AjamianL, GallouziIE, BernardN, MoulandAJ . EEF2 and Ras- GAP SH3 domain-binding protein (G3BP1) modulate stress granule assembly during HIV-1 infection Nat Commun, 2014,5:4819. [本文引用: 1]
JiménezVC, MartinezFO, BooimanT, van DortKA, van de KlundertMAA, GordonS, GeijtenbeekTB, KootstraNA . G3BP1 restricts HIV-1 replication in macrophages and T-cells by sequestering viral RNA Virology, 2015,486:94-104. URL [本文引用: 1]
CintiA, Le SageV, GhanemM, MoulandAJ . HIV-1 gag blocks selenite-induced stress granule assembly by altering the mRNA Cap-Binding complex MBio, 2016,7(2):e00329. [本文引用: 1]
RaoS, TemziA, AmorimR, YouJC, MoulandAJ . HIV-1 NC-induced stress granule assembly and translation arrest are inhibited by the dsRNA binding protein Staufen1 RNA, 2018,24(2):219-236. URL [本文引用: 1]
NelsonEV, SchmidtKM, DeflubéLR, Do?anayS, BanadygaL, OlejnikJ, HumeAJ, RyabchikovaE, EbiharaH, KedershaN, HaT, MühlbergerE . Ebola virus does not induce stress granule formation during infection and sequesters stress granule proteins within viral inclusions J Virol, 2016,90(16):7268-7284. URL [本文引用: 1]
LuB, NakamuraT, InouyeK, LiJ, TangY, Lundb?ckP, Valdes-FerrerSI, OlofssonPS, KalbT, RothJ, ZouY, Erlandsson-HarrisH, YangH, TingJP, WangH, AnderssonU, AntoineDJ, ChavanSS, HotamisligilGS, TraceyKJ . Novel role of PKR in inflammasome activation and HMGB1 release Nature, 2012,488(7413):670-674. [本文引用: 1]
TaghaviN, SamuelCE . Protein kinase PKR catalytic activity is required for the PKR-dependent activation of mitogen-activated protein kinases and amplification of interferon beta induction following virus infection Virology, 2012,427(2):208-216. URL [本文引用: 1]
ReikineS, NguyenJB, ModisY . Pattern recognition and signaling mechanisms of RIG-I and MDA5 Front Immunol, 2014,5:342. [本文引用: 1]
WuB, HurS . How RIG-I like receptors activate MAVS Curr Opin Virol, 2015,12:91-98. URL [本文引用: 1]
SilvermanRH . Viral encounters with 2°,5°-oligoadenylate synthetase and RNase L during the interferon antiviral response J Virol, 2007,81(23):12720-12729. URL [本文引用: 1]
ChaudhryY, NayakA, BordeleauME, TanakaJ, PelletierJ, BelshamGJ, RobertsLO, GoodfellowIG . Caliciviruses differ in their functional requirements for eIF4F components J Biol Chem, 2006,281(35):25315-25325. URL [本文引用: 1]
ClavarinoG, CláudioN, DaletA, TerawakiS, CoudercT, ChassonL, CeppiM, SchmidtEK, WengerT, LecuitM, GattiE, PierreP . Protein phosphatase 1 subunit Ppp1r15a/ GADD34 regulates cytokine production in polyinosinic: polycytidylic acid-stimulated dendritic cells Proc Natl Acad Sci USA, 2012,109(8):3006-3011. URL [本文引用: 1]
 ,中国医学科学院/北京协和医学院病原生物学研究所,国家卫生健康委员会病原系统生物学重点实验室,北京 100730
,中国医学科学院/北京协和医学院病原生物学研究所,国家卫生健康委员会病原系统生物学重点实验室,北京 100730
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}