关键词:谷子; 核心种质; 表型性状; 综合评价 Phenotypic Diversity Evaluations of Foxtail Millet Core Collections WANG Hai-Gang1, JIA Guan-Qing2, ZHI Hui2, WEN Qi-Fen1, DONG Jun-Li1, CHEN Ling1, WANG Jun-Jie1, CAO Xiao-Ning1, LIU Si-Chen1, WANG Lun1, QIAO Zhi-Jun1,*, DIAO Xian-Min2,* 1 Institute of Crop Germplasm Resources, Shanxi Academy of Agricultural Sciences / Key Laboratory of Crop Gene Resources and Germplasm Enhancement on Loess Plateau, Ministry of Agriculture / Shanxi Key Laboratory of Genetic Resources and Genetic Improvement of Minor Crops, Taiyuan 030031, China
2Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
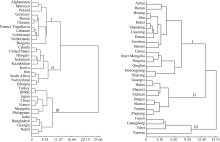
Fund:This study was supported by the National Natural Science Foundation of China (31301328, 31171560), the National Key Technology Research and Development Program (2014BAD07B01), the Special Program of Modern Agro-industry Technology System (CARS07-12.5-A02, CARS-07-12.5-A12) and the Agricultural Science and Technology Innovation Program of CAAS AbstractEvaluation of genetic diversity could benefit the identification of optimal parental combinations for obtaining segregating offspring with maximum genetic variability, and facilitate the introgression of favorable genes from various germplasm into commercial cultivars. In this study, foxtail millet core collections including 878 world-wide accessions were evaluated through phenotypic analysis of 15 agronomic traits. Main conclusions are as follows: (1) Chinese accessions present higher level of phenotypic diversity, especially in grain weight per main stem, panicle length, panicle diameter, plant height, stem node number and growth period; phenotypic diversity is lower in elite cultivars than in traditional landraces; gains of breeding progress in foxtail millet are expressed in reduced plant height and panicle length, coupling with increased diameter of main stem, stem node number, panicle diameter, grain weight per main stem, panicle weight per main stem and growth period; (2) from the cluster analysis, foreign resources are divided into three categories according to the geographical origins, the first category mainly includes accessions from northeast European countries, the second cluster is mainly collected from North America and Africa, and the third group includes varieties mainly from East Asia, South Asia, African, and European countries; Chinese collections is able to be divided into three groups of spring-sowing, summer-sowing and southern ecotypes; (3) comprehensive assessment of phenotypic traits based on principal component analysis (PCA) and step regression analysis demonstrates that leaf sheath color, bristle length, hull color, grain color, plant height, spike length, stem diameter and grain weight per plant can be used as main identification indicators of foxtail millet phenotypic variations. This research will benefit the utilization of foxtail millet resources in variety breeding practices in the future.
Keyword:Foxtail millet; Core collections; Phenotypic traits; Comprehensive evaluations Show Figures Show Figures
4 结论建立了回归方程, 筛选出8个性状作为谷子表型鉴定的主要指标, 能有效反映谷子种质的综合表现。878份谷子的应用核心种质变异丰富, 在遗传育种中有较好的利用价值。我国谷子的育种水平还较低, 育种选择和进步的空间还很大。山西省适合谷子表型精准鉴定, 鉴定数据全面稳定且可为应用核心种质在山西的应用以及其他谷子产区的育种提供参考。 The authors have declared that no competing interests exist.
刁现民. 中国谷子产业与产业技术体系. 北京: 中国农业科学与技术出版社, 2011. pp 20-30Diao XM. Chinese Industry and Technical System of Foxtail Millet. Beijing: China Agricultural Sciences and Technology Press, 2011. pp 20-30(in Chinese)[本文引用:1]
[2]
智慧, 牛振刚, 贾冠清, 柴杨, 李伟, 王永芳, 李海权, 陆平, 白素兰, 刁现民. 谷子干草饲用品质性状变异及相关性分析. 作物学报, 2012, 38: 800-807ZhiH, Niu ZG, Jia GQ, ChaiY, LiW, Wang YF, Li HQ, LuP, Bai SL, Diao XM. Variation and correlation analysis of hay forage quality traits of foxtail millet [Setaria italica (L. ) Beauv. ]. Acta Agron Sin, 2012, 38: 800-807 (in Chinese with English abstract)[本文引用:1]
[3]
刘三才, 朱志华, 李为喜, 刘方, 李燕, 黄蓉. 谷子品种资源微量元素硒和蛋白质含量的测定与评价. 中国农业科学, 2009, 42: 3812-3818Liu SC, Zhu ZH, Li WX, LiuF, LiY, HuangR. Evaluation of selenium and protein content of foxtail millet land races originated from different ecological regions of China. Sci Agric Sin, 2009, 42: 3812-3818 (in Chinese with English abstract)[本文引用:2]
[4]
刘敏轩, 陆平. 中国谷子育成品种维生素E含量分布规律及其与主要农艺性状和类胡萝卜素的相关性分析. 作物学报, 2013, 39: 398-408Liu MX, LuP. Distribution of vitamin E content and its correlation with agronomic trait sand carotenoids content in foxtail millet varieties in china. China. Acta Agron Sin, 2013, 39: 398-408 (in Chinese with English abstract)[本文引用:2]
[5]
邵丽华, 王莉, 白文文, 刘雅娟. 山西谷子资源叶酸含量分析及评价. 中国农业科学, 2014, 47: 1265-1272Shao LH, WangL, Bai WW, Liu YJ. Evaluation and Analysis of the folic acid content in millet from the different ecological regions in Shanxi province. Sci Agric Sin, 2014, 47: 1265-1272 (in Chinese with English abstract)[本文引用:2]
[6]
Yang XY, Wan ZW, PerrycL, Lue HY, WangQ, Zhao CH, LiJ, XieF, Yu JC, Cui TX, WangT, Li MQ, Ge QS. Early millet use in northern China. Proc Natl Acad Sci USA, 2012, 109: 3726-3730. [本文引用:1]
[7]
Jia GQ, Huang XH, ZhiH, ZhaoY, ZhaoQ, Li WJ, ChaiY, Yang LF, Liu KY, Lu HY, Zhu CR, Lu YQ, Zhou CC, Fan DL, Weng QJ, Guo YL, HuangT, ZhangL, Lu TT, FengQ, Hao HF, Liu HK, LuP, ZhangN, Li YH, Guo EH, Wang SJ, Wang SY, Liu JR, Zhang WF, Chen GQ, Zhang BJ, LiW, Wang YF, Li HQ, Zhao BH, Li JY, Diao XM, HanB. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (Setaria italica). Nat Genet, 2013, 45: 957-961[本文引用:3]
[8]
Frankel OH. Genetic perspectives of germplasm conservation. In: Arber W, Llimensee K, Peacock W J, Starlinger P, eds. Genetic Manipulation: Impact on Man and Society. Cambridge, UK: Cambridge University Press, 1984. pp 161-170[本文引用:1]
[9]
李自超, 张洪亮, 曹永生, 裘宗恩, 魏兴华, 汤圣祥, 余萍, 王象坤. 中国地方稻种资源初级核心种质取样策略研究. 作物学报, 2003, 29: 20-24Li ZC, Zhang HL, Cao YS, Qiu ZE, Wei XH, Tang SX, YuP, Wang XK. Studies on the sampling strategy for primary core collection of Chinese ingenious rice. Acta Agron Sin, 2003, 29: 20-24 (in Chinese with English abstract)[本文引用:1]
[10]
胡标林, 万勇, 李霞, 雷建国, 罗向东, 严文贵, 谢建坤. 水稻核心种质表型性状遗传多样性分析及综合评价. 作物学报, 2012, 38: 829-839Hu BL, WanY, LiX, Lei JG, Luo XD, Yan WG, Xie JK. Analysis on genetic diversity of phenotypic traits in rice (Oryza sativa) core collection and its comprehensive assessment. Acta Agron Sin, 2012, 38: 829-839 (in Chinese with English abstract)[本文引用:3]
[11]
郝晨阳, 董玉琛, 王兰芬, 游光霞, 张洪娜, 盖红梅, 贾继增, 张学勇. 我国普通小麦核心种质的构建及遗传多样性分析. 科学通报, 2008, 53: 908-915Hao CY, Dong YC, Wang LF, You GX, Zhang HN, Ge HM, Jia JZ, Zhang XY. Genetic diversity analysis and construction core germplasm of common wheat in china. Chin Sci Bull, 2008, 53: 908-915 (in Chinese with English abstract)[本文引用:1]
[12]
魏添梅, 昌小平, 闵东红, 景蕊莲. 小麦抗旱品种的遗传多样性分析及株高优异等位变异挖掘. 作物学报, 2010, 36: 895-904Wei TM, Chang XP, Min DH, Jing RL. Analysis of genetic diversity and tapping elite alleles for plant height in drought- tolerant wheat varieties. Acta Agron Sin, 2010, 36: 895-904 (in Chinese with English abstract)[本文引用:1]
[13]
刘志斋. 中国玉米地方品种的多样性研究及种族划分. 西南大学博士学位论文, 重庆, 2008Liu ZZ. Diversity Analysis and Racial Classification of Maize Land races in China. PhD Dissertation of Southwest University, Chongqing, China (in Chinese with English abstract)[本文引用:1]
[14]
姚坚强, 鲍坚东, 朱金庆, 桂毅杰, 沈秋芳, 胡伟民, 樊龙江. 中国糯玉米wx基因种质资源遗传多样性. 作物学报, 2013, 39: 43-49Yao JQ, Bao JD, Zhu JQ, Gui YJ, Shen QF, Hu WM, Fan LJ. Genetic diversity of waxy gene in Chinese glutinous maize. Acta Agron Sin, 2013, 39: 43-49 (in Chinese with English abstract)[本文引用:1]
[15]
宋喜娥, 李英慧, 常汝镇, 郭平毅, 邱丽娟. 中国栽培大豆(Glycine max (L. ) Merr. )微核心种质的群体结构与遗传多样性. 中国农业科学, 2010, 11: 2209-2219Song XE, Li YH, Chang RZ, Guo PY, Qiu LJ. Population structure and genetic diversity of mini core collection of cultivated soybean (Glycine max (L. ) Merr. ) in China. Sci Agric Sin, 2010, 43: 2209-2219 (in Chinese with English abstract)[本文引用:1]
[16]
文自翔, 赵团结, 丁艳来, 盖钧镒. 中国栽培及野生大豆的遗传多样性、地理分化和演化关系研究. 科学通报, 2009, 54: 3301-3310Wen ZX, Zhao TJ, Ding YL, Gai JY. Study on the relationship between cultivated and wild soybean variety in china: Genetic diversity, geographical differentiation and evolutionary relationship. Chin Sci Bull, 2009, 54: 3301-3310 (in Chinese with English abstract)[本文引用:1]
[17]
陈雨, 潘大建, 杨庆文, 刘斌, 范芝兰, 陈建酉, 李晨. 广东高州野生稻应用核心种质取样策略, 作物学报, 2009, 35: 459-466 (in Chinese with English abstract)ChenY, Pan DJ, Yang QWen, LiuB, Fan ZL, Chen JY, LiC. Sampling strategy for an applied core collection of gaozhou wild rice (Oryza rufipogon Griff. ) in Guangdong, China. Acta Agron Sin, 2009, 35: 459-466[本文引用:1]
[18]
Diao XM, SchnableJ, Bennetzen JL, Li JY. Initiation of Setaria as a model plant. Front Agric Sci Eng, 2014, 1: 16-20. [本文引用:1]
[19]
范志影, 陆平, 刘方, 张萍, 李为喜, 朱志华. 谷子初级核心种质生育酚的组分及其评价. 植物遗传资源学报, 2009, 10: 378-384Fan ZY, LuP, LiuF, ZhangP, Li WX, Zhu ZH. Identification and evalution of tocopherol content in primary core-collection of foxtail millet. J Plant Genet Resour, 2009, 10: 378-384 (in Chinese with English abstract)[本文引用:1]
[20]
田伯红. 谷子地方品种和育成品种的遗传多样性研究. 植物遗传资源学报. 2010, 11: 224-228Tian BH. Genetic diversity of land race and improved cultivars in foxtail millet. J Plant Genet Resour, 2010, 11: 224-228 (in Chinese with English abstract)[本文引用:2]
[21]
王晓娟, 祁旭升, 王兴荣, 苏俊阳. 甘肃省谷子地方种质资源遗传多样性分析. 干旱地区农业研究, 2009, 27(6): 129-153Wang XJ, Qi XS, Wang XM, Su JY. Genetic diversity of foxtail land race germplasm in Gansu Province. Agric Res Arid Areas, 2009, 27(6): 129-153 (in Chinese with English abstract)[本文引用:2]
[22]
吴舒致, 黎裕. 谷子种质资源的主成分分析和图论主成分分类. 西北农业学报, 1997, 6(2): 46-50Wu SZ, LiY. Principal component analysis and graph theory. Acta Agric Boreali-Occid Sin, 1997, 6(2): 46-50 (in Chinese with English abstract)[本文引用:1]
[23]
王节之, 郝晓芬, 王根全, 王潞英, 孙美荣. 谷子种质资源分子标记的多态性研究. 生物技术, 2006, 16(1): 10-14Wang JZ, Hao XF, Wang GQ, Wang LY, Sun MR. Study of genetic diversity of millet germplasm resources by molecular markers. Biotechnology, 2006, 16(1): 10-14 (in Chinese with English abstract)[本文引用:1]
[24]
朱学海, 张艳红, 宋燕春, 赵治海, 刘志斋, 石云素, 黎裕, 王天宇. 基于SSR标记的谷子遗传多样性研究. 植物遗传资源学, 2010, 11: 698-702Zhu XH, Zhang YH, Song YC, Zhao ZH, Liu ZZ, Shi YS, LiY, Wang TY. Genetic diversity analysis of foxtail millet accessions revealed by SSR markers. J Plant Genet Resour, 2010, 11: 698-702 (in Chinese with English abstract)[本文引用:1]
[25]
贾小平, 谭贤杰, 李永祥, 王天宇, 黎裕. 用SSR标记研究谷子品种的遗传多样性. 江西农业大学学报, 2009, 31: 633-638Jia XP, Tan XJ, Li YX, Wang TY, LiY. A study on the genetic diversity of foxtail millet cultivars by SSR markers. Acta Agic Univ Jiangxiensis, 2009, 31: 633-638 (in Chinese with English abstract)[本文引用:1]
[26]
孙加梅, 王雪梅, 王东建, 杨延兵, 李华, 段丽丽, 王晓宇, 张晗. 谷子种质资源遗传多样性研究. 山东农业科学, 2013, 45(3): 33-37Sun JM, Wang XM, Wang DJ, Yang YB, LiH, Duan LL, Wang XY, ZhangH. Analysis genetic diversity of foxtail millet germplasm resources. Sci Agric Shand ong, 2013, 45(3): 33-37 (in Chinese with English abstract)[本文引用:2]
[27]
杨天育, 黄相国, 何继红, 沈裕琥, 吴国忠. 谷子遗传资源多样性研究进展. 西北农业学报, 2003, 12(1): 43-47Yang TY, Huang XG, He JH, Shen YH, Wu GZ. Advance in diversity of genetic resource of foxtail millet. Acta Agric Boreali-occident Sin, 2003, 12(1): 43-47 (in Chinese with English abstract)[本文引用:2]
[28]
孟庆立, 关周博, 冯佰利, 柴岩, 胡银岗. 谷子抗旱相关性状的主成分与模糊聚类分析. 中国农业科学, 2009, 42: 2667-2675Meng QL, Guan ZB, Feng BL, ChaiY, Hu YG. Principal component analysis and fuzzy clustering on drought tolerance related traits of foxtail millet (Setaria italica). Sci Agric Sin, 2009, 42: 2667-2675 (in Chinese with English abstract)[本文引用:1]
[29]
温琪汾, 刘润堂, 王纶, 王星玉, 师颖. 谷子种质资源抗黑穗病鉴定与过氧化物酶研究. 植物遗传资源学报, 2006, 7: 349-351Wen QF, Liu RT, WangL, Wang XY, ShiY. Screening of foxtail millet germplasm for resistance to grain smut and related peroxidase study. J Plant Genet Resour, 2006, 7: 349-351 (in Chinese with English abstract)[本文引用:1]
[30]
陆平. 谷子种质资源描述规范和数据标准. 北京: 中国农业出版社, 2006LuP. Resource Description Specification and Date Stand ard of Foxtail Millet Germplasm. Beijing: China Agriculture Press, 2006 (in Chinese)[本文引用:1]
[31]
Kim EJ, Sa KJ, Park KC, Lee JK. Study of genetic diversity and relationships among accessions of foxtail millet [Setaria italica (L. ) P. Beauv. ] in Korea, China, and Pakistan using SSR markers. Genes Genom, 2012, 34: 529-538[本文引用:1]
[32]
Lin HS, Liao GI, Chiang CY, Kuoh CS, Chang SB. Genetic diversity in the foxtail millet (Setaria italica) germplasm as determined by agronomic traits and microsatellite markers. Aust J Crop Sci, 2012, 6: 342-349[本文引用:1]
[33]
罗俊杰, 欧巧明, 叶春雷, 王方, 王镛臻, 陈玉梁. 重要胡麻栽培品种的抗旱性综合评价及指标筛选, 作物学报, 2014, 40: 1259-1273 (in Chinese with English abstract)Luo JJ, Ou QM, Ye CL, WangF, Wang YZ, Chen YL. Comprehensive valuation of drought resistance and screening of indices of important flax cultivars. Acta Agron Sin, 2014, 40: 1259-1273[本文引用:1]
[34]
王海岗, 陈凌, 王君杰, 曹晓宁, 董俊丽, 王纶, 杨天育, 乔治军. 20份山西糜子种质资源抗旱性综合评价. 中国农学通报, 2014, 30(36): 115-119Wang HG, ChenL, Wang JJ, Cao XN, Dong JL, WangL, Yang TY, Qiao ZJ. Comprehensive assessment of drought resistance of 20 proso millet germplasm resources in whole growth period. Chin Agric Sci Bull, 2014, 30(36): 115-119[本文引用:1]
[35]
Wang CF, Jia GQ, ZhiH, Niu ZG, ChaiY, LiW, Wang YF, Li HQ, LuP, Zhao BH, Diao XM. Genetic diversity and population structure of Chinese foxtail millet [Setaria italica (L. ) Beauv. ] land races. Genes Genom Genet, 2012, 2: 769-777[本文引用:1]
[36]
Jia GQ, Liu XT, Schnable JC, Niu ZG, Wang CF, Li YH, Wang SJ, Wang SY, Liu JR, Guo EH, ZhiH, Diao XM. Microsatellite variations of elite setaria varieties released during last six decades in China. PLoS One, 2015, 10(5): e0125688. doi: 10.1371/journal.pone.0125688[本文引用:1]
[37]
王殿赢, 陈玉香. 谷子品种南北异地种植主要性状变化及其应用. 中国农业科学, 1980, (4): 33-38Wang DY, Chen YX. The main characters variation of the miller varieties grown both in north and south areas and its use in practice, Sci Agric Sin, 1980, (4): 33-38 (in Chinese with English abstract)[本文引用:1]
[38]
古兆明, 古世禄. 山西谷子起源与发展研究. 北京: 中国农业科学技术出版社, 2007. pp 3-7Gu ZM, Gu SL. Origin and development of Shanxi foxtail millet. Beijing. Agricultural Science and Technology Press, 2007. pp 7-10(in Chinese)[本文引用:1]
 , 刁现民
, 刁现民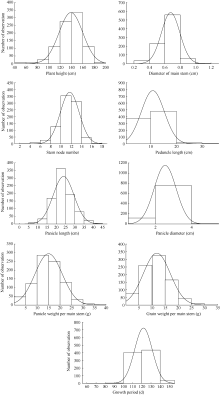



{kind=link}
{kind=link}
{kind=link}