关键词:遗传背景; 稻瘟病菌; 致病性; 水稻单基因系; 基因聚合 Comparison of Pathogenicity of Pyricularia oryzaeunder Different Genetic Backgrounds MA Jun-Tao1, ZHANG Guo-Min1,*, XIN Ai-Hua1, ZHANG Li-Yan1, DENG Ling-Wei1, WANG Yong-Li1, WANG Ying1, REN Yang1, GONG Xiu-Jie1, GE Xuan-Liang1, YANG Xiu-Feng2 1Crop Cultivation Research Institute, Heilongjiang Academy of Agricultural Sciences, Harbin 150086, China
2 The Heihe Branch of Heilongjiang Academy of Agricultural Science, Heihe 164300, China
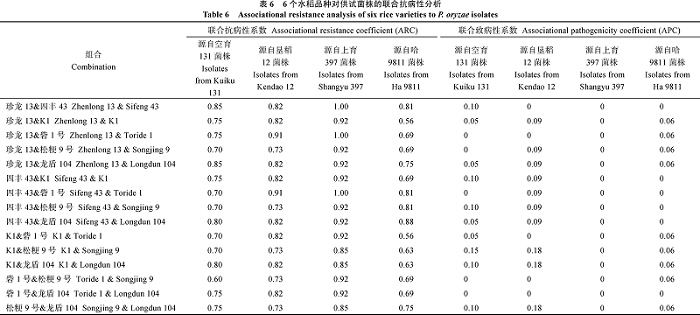
Fund:This study was supported by Science Foundation for Distinguished Young Scholars of Heillongjiang Province (JC201214) and the Agricultural Science and Technology Innovation Program of Heillongjiang Province (2013ZD019) AbstractRice blast is one of the most serious rice diseases in Heilongjiang province, resulting in high incidence and yield reduction. In order to definite the pathogenicity of P. oryzaeisolates and resistance improvement of nurse varieties under different genetic backgrounds, the experiment was conducted using four groups of Pyricularia oryzae ( P. oryzae) isolates and rice varieties including Chinese differential varieties, Japanese differential varieties, monogenic rice lines and landrace. Using the method of spraying inoculation and associational resistance and gene pyramiding analyses, the main results were as follows: (1) Under the Chinese differential varieties system, the P. oryzaeisolates from Kuiku 131, Kendao 12, Shangyu 397, and Ha 9811 were divided into ten, six, six, and nine physiological races respectively, and predominant physiological race of P. oryzaeisolates from Ha 9811 was ZF1 while the predominant physiological races of the other three groups of P. oryzaeisolates were all ZG1. (2) Under the Japanese differential varieties system, the P. oryzaeisolates from Kuiku 131, Kendao 12, Shangyu 397, and Ha 9811 were divided into 19, 11, 13, and 14 physiological races respectively, furthermore, the average disease incidence was 54.17%, 46.21%, 41.67%, and 59.90%, respectively. (3) Under the monogenic rice lines system, the P. oryzaeisolates from Kuiku 131, Kendao 12, Shangyu 397 and Ha 9811 were divided into 20, 11, 12, and 16 pathogenic types respectively, additionally, their similarity coefficient of pathogenicity was 0.20-0.87, 0.25-0.90, 0.25-1.00, and 0.31-0.88, respectively. (4) Under the landrace system, the P. oryzaeisolates from Kuiku 131, Kendao 12, Shangyu 397, and Ha 9811 were divided intotwelve, sever, nine and thirteen pathogenic types respectively, with the best blast-resistance varieties of Longdun 104, Songjing 6, Songjing 9, and Longdao 7. (5) Using the method of associational resistance, the average associational resistance coefficient and associational pathogenicity coefficient were 0.71-0.92 and 0-0.08, respectively; Using the method of gene pyramiding, the number of immunity gene combination to the P. oryzaeisolates from Kuiku 131, Kendao 12, Shangyu 397, and Ha 9811 were two, two, five and seven. In conclusion, using monogenic rice lines to research the pathogenicity of P. oryzae is better than using other appraisal systems. And good effects can be get from two resistance improvement methods, but both of them have their advantages and disadvantages.
Keyword:Genetic background; Pyricularia oryzae; Pathogenicity; Monogenic rice lines; Gene pyramiding Show Figures Show Figures
张国民, 马军韬, 肖佳雷, 刘迎雪, 辛爱华, 任洋, 张丽艳, 刘东风. 已知抗瘟基因在黑龙江省寒地稻区的评价与利用. 植物病理学报, 2011, 41: 72-79Zhang GM, Ma JT, Xiao JL, Liu YX, Xin AH, RenY, Zhang LY, Liu DF. Evaluation and utilization of value of twenty-four blast resistance genes in north cold region, Heilongjiang. Acta Phytopathol Sin, 2011, 41: 72-79 (in Chinese with English abstract)[本文引用:1]
[2]
雷财林, 张国民, 程治军, 马军滔, 王久林, 辛爱华, 陈平, 肖家雷, 张欣, 刘迎雪, 郭秀平, 王洁, 翟虎渠, 万建民. 黑龙江省稻瘟病菌生理小种毒力基因分析及抗病育种策略. 作物学报, 2011, 37: 18-27Lei CL, Zhang GM, Cheng ZZ, Ma JT, Wang JL, Xin AH, ChenP, Xiao JL, ZhangX, Liu YX, Guo XP, WangJ, Zhai HQ, Wan JM. Pathogenic races and virulence gene structure of Magnaporthe oryzae population and rice breeding strategy blast resistance in Heilongjiang province. Acta Agron Sin, 2011, 37: 18-27 (in Chinese with English abstract)[本文引用:1]
[3]
Flor HH. Current status of the gene for gene concept. Annu Rev Phytopathol, 1971, 9: 275-296[本文引用:1]
[4]
全国稻瘟病菌生理小种联合试验组. 我国稻瘟病菌生理小种研究. 植物病理学报, 1980, 10: 71-81China Coorporation of Research on Physiological Races of Pyricularia oryzae. Research on physiological races of rice blast fungus in China. Acta Phytopathol Sin, 1980, 10: 71-81 (in Chinese with English abstract)[本文引用:1]
[5]
KiyosawaS. Gene analysis for blast resistance. Oryzae, 1981, 18: 196-203[本文引用:1]
[6]
HiroshiT, Mary J T Y, Ebron L A, Hayashi N, Ando I, Kato H, Imbe T, Khush G S. Development of monogenic lines of rice for blast resistance. Breed Sci, 2000, 50: 229-234[本文引用:1]
[7]
刘志恒, 王世维, 魏松红, 王海宁, 郑文静, 杨廷章. 2011-2012年辽宁省稻瘟病菌种群动态分析. 沈阳农业大学学报, 2014, 45: 393-397Liu ZH, Wang SW, Wei SH, Wang HN, Zheng WJ, Yang TZ. Dynamics of rice blast fungus of population in Liaoning Province during in 2011-2012. J Shenyang Agric Univ, 2014, 45: 393-397 (in Chinese with English abstract)[本文引用:2]
[8]
马军韬, 张国民, 辛爱华, 肖佳雷, 刘迎雪, 宫秀杰. 黑龙江省稻瘟病菌生理小种鉴定与分析. 植物保护, 2010, 36(3): 97-99Ma JT, Zhang GM, Xin AH, Xiao JL, Liu YX, Gong XJ. Identification and analysis of the physiologic races of Magnaporthe grisea in Heilongjiang province. Plant Prot, 2010, 36(3): 97-99 (in Chinese with English abstract)[本文引用:1]
[9]
杨秀娟, 阮宏椿, 杜宜新, 陈福如, 王茂明. 福建省稻瘟病菌致病性及其无毒基因分析. 植物保护学报, 2007, 34: 337-342Yang XJ, Ruan HC, Du YX, Chen FR, Wang MM. Pathogenicity and avirulence genes analysis of Magnaporthe grisea Barr from rice in Fujian province of China. Acta Phytophyl Sin, 2007, 34: 337-342 (in Chinese with English abstract)[本文引用:2]
[10]
肖丹凤, 王玲, 刘连盟, 侯恩庆, 黄世文. 黑浙桂稻瘟病菌生理小种鉴定与遗传多样性分析. 西南农业学报, 2014, 27: 121-126Xiao DF, WangL, Liu LM, Hou EQ, Huang SW. Physiological races identification and genetic diversity analysis of Magnaporthe grisea in Province of Heilongjiang, Zhejiang and Guangxi. Southwest China J Agric Sci, 2014, 27: 121-126 (in Chinese with English abstract)[本文引用:1]
[11]
刘文德, 阮志平, 郑士琴, 赵长江, 郑武, 鲁国东, 王忠华. 水稻主要抗瘟基因对福建稻瘟菌群体的抗性分析. 植物病理学报, 2005, 35: 526-531Liu WD, Ruan ZP, Zheng SQ, Zhao CJ, ZhengW, Lu GD, Wang ZH. Resistance of rice major Pi-genes to the Magnaporthe grisea population in Fujian, China. Acta Phytopathol Sin, 2005, 35: 526-531 (in Chinese with English abstract)[本文引用:1]
[12]
李湘民, 兰波, 黄凌洪, 何烈干, 张天才, 黄瑞荣. 江西省稻瘟病菌的致病性分化. 植物保护学报, 2009, 36: 497-503Li XM, LanB, Huang LH, He LG, Zhang TC, Huang RR. Pathogenicity differentiation of Magnaporthe grisea (Hebert) Barr. from rice in Jiangxi province of China. Acta Phytophyl Sin, 2009, 36: 497-503 (in Chinese with English abstract)[本文引用:1]
[13]
宋成艳, 王桂玲, 辛爱华, 丛万彪. 黑龙江省水稻品种空育131稻瘟病菌生理小种种类及发病原因分析. 黑龙江农业科学, 2007, (1): 41-42Song CY, Wang GL, Xin AH, Cong WB. Analysis on kinds of rice blast races in Kongyu 131 and its reasons of pathologic reaction. Heilongjiang Agric Sci, 2007, (1): 41-42 (in Chinese with English abstract)[本文引用:1]
[14]
兰波, 杨迎青, 常冬冬, 徐沛东, 李湘民. 基因丽江新团黑谷的稻瘟病菌致病性分化. 华中农业大学学报, 2015, 34(1): 28-32LanB, Yang YQ, Chang DD, Xu PD, Li XM. Pathogenicity differentiation of rice blast pathogen (Magnaporthe grisea) based on Lijiangxintuanheigu. J Huazhong Agric Univ, 2015, 34(1): 28-32 (in Chinese with English abstract)[本文引用:2]
[15]
周江鸿, 王久林, 蒋琬如, 雷财林, 凌忠专. 我国稻瘟病菌毒力基因的组成及其地理分布. 作物学报, 2003, 29: 646-651Zhou JH, Wang JL, Jiang WR, Lei CL, Ling ZZ. Virulence genes diversity and geographic distribution of Pyricularia grisea in China. Acta Agron Sin, 2003, 29: 646-651 (in Chinese with English abstract)[本文引用:3]
[16]
Bonman JM, Vergel de Dios T I, Khin M M. Physiologic specialization of Pyricularia oryzae in the Philippines. Plant Dis, 1986, 70: 767-769[本文引用:1]
[17]
Mackill DJ, Bonman JM. Inheritance of blast resistance in near-isogenic lines of rice. Phytopathology, 1992, 82: 746-749[本文引用:1]
马辉刚, 曹九龙, 胡水秀, 黄瑞荣, 涂雪琴. 水稻品种对稻瘟病的抗性分析和利用评价. 江西农业大学学报, 2006, 28: 213-216Ma HG, Cao JL, Hu XS, Huang RR, Tu XQ. Identification and evaluation of resistance of rice to Magnaporthe grisea. Acta Agric Univ Jiangxiensis, 2006, 28: 213-216 (in Chinese with English abstract)[本文引用:1]
[20]
袁洁, 杨学辉, 何海永. 部分水稻品种对稻瘟病群体的抗病性分析. 种子, 2006, 25(8): 13-16YuanJ, Yang XH, He HY. Analysis of resistance to Magnaporthe grisea populations in some rice varieties. Seed, 2006, 25(8): 13-16 (in Chinese with English abstract)[本文引用:1]
[21]
Liu XQ, Yang QZ, LiuF, Hua LX, Wang CT, Pan QH. Identification and fine mapping of Pi-39(t), a major gene coffering the board-spectrum resistance to Magnaporthe oryzae. Mol Genet Genomics, 2007, 248: 403-410[本文引用:1]
[22]
LeeS, CostanzoS, JiaY, Olsen KM, Caicedo AL. Evolutionary dynamics of the genomic region around the blast resistance gene Pi-ta in AA genome Oryza species. Genetics, 2009, 183: 1315-1325[本文引用:1]
[23]
Xiao WM, Yang QY, WangH, GuoT, Liu YZ, Zhu XY, Chen ZZ. Identification and fine mapping of a resistance gene to Magnaporthe oryzae in a space-induced rice mutant. Mol Breed, 2011, 28: 303-312[本文引用:1]
[24]
YuanB, ZhaiC, Wang WJ, Zeng XS, Xu XK, Hu HQ, LinF, WangL, Pan QH. The Pik-p resistance to Magnaporthe oryzae in rice is mediated by a pair of closely linked CC-NBS-LRR gene. Theor Appl Genet, 2011, 122: 1017-1028[本文引用:1]
[25]
Zhu XY, ChenS, Yang JY, Zhou SC, Zeng LX, HanJ, SuJ, WangL, Pan QH. The identification of Pi50(t), a new member of rice blast resistance Pi2/Pi9 multigene family. Theor Appl Genet, 2012, 124: 1295-1304[本文引用:1]
[26]
JiangH, Feng YT, BaoL, LiX, Gao GJ, Zhang QL, Xiao JH, Xu CG, He YQ. Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding. Mol Breed, 2012, 30: 1679-1688[本文引用:1]
[27]
Xiao WM, Yang QY, WangH, DuanJ, GuoT, Liu YZ, Zhu XY, Chen ZQ. Identification and fine mapping of a major R gene to Magnaporthe oryzae in a broad-spectrum resistant germplasm in rice. Mol Breed, 2012, 30: 1715-1726[本文引用:1]
[28]
WangY, WangD, Deng XJ, LiuJ, SunP, LiuY, HuangH, JiangN, KangH, NingY, WangZ, XiaoY, LiuX, LiuE, DaiL, Wang GL. Molecular mapping of the blast resistance genes Pi2-1 and Pi51(t) in the durably resistant rice “Tianjingyeshengdao”. Phytopathology, 2012, 102: 779-786[本文引用:1]
[29]
JiangN, Li ZQ, WuJ, WangY, Wu LJ, Wang SH, WenT, LiangY, Sun PY, Liu JL, Dai LY, Wang ZL, WangC, Luo MZ, Liu XL, Wang GL, WangD. Molecular mapping of the Pi2/9 allelic gene Pi2-2 conferring broad-spectrum resistance to Magnaporthe oryzae in the rice cultivar Jefferson. Rice, 2012, 5: 29[本文引用:1]
[30]
LiuY, LiuB, Zhu XY, Yang JY, AliciaB, Wang GL, Jan EL, HeiL, LiuY. Fine-mapping and molecular marker development for Pi56(t), a NBS-LRR gene conferring broad-spectrum resistance go Magnaporthe oryzae in rice. Theor Appl Genet, 2013, 126: 985-998[本文引用:1]
 , 辛爱华
, 辛爱华