关键词:小麦籽粒性状; TaGW2基因; 等位变异; 高分辨率熔解曲线(HRM); 分子标记辅助选择 Composition and Selection of TaGW2-6A Alleles for Wheat Kernel Weight KOU Cheng, GAO Xin, LI Li-Qun, LI Yang, WANG Zhong-Hua, LI Xue-Jun* College of Agronomy, Northwest A&F University / State Key Laboratory of Crop Stress Biology in Arid Areas, Yangling 712100, China
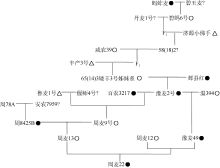
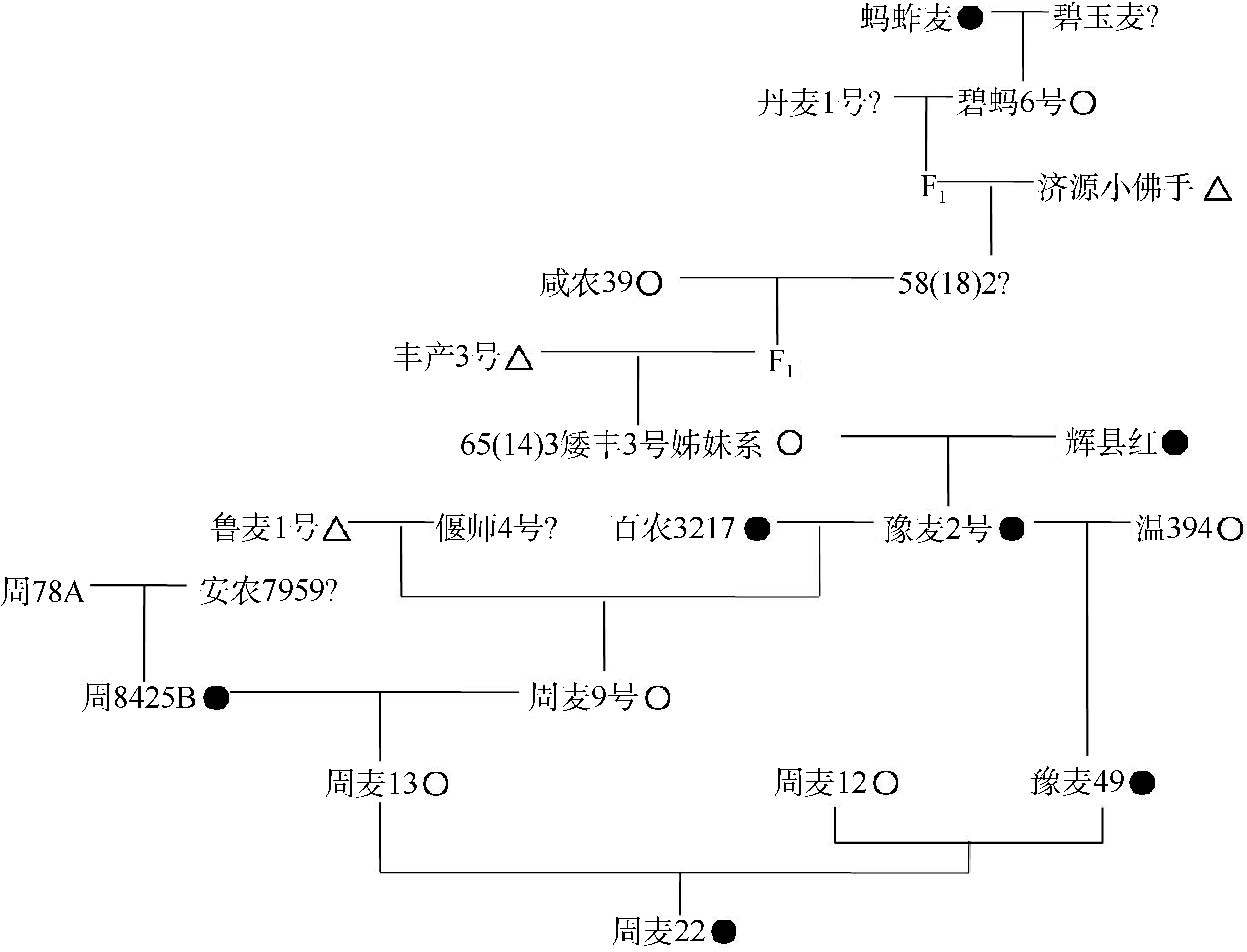
Abstract TaGW2 on chromosome 6A is a key gene governing kernel size of wheat ( Triticum aestivumL.). There are a single T-base insertion in the eighth exon of TaGW2 (977 bp) and two SNPs (Hap-6A-A and Hap-6A-G) in the promoter region. In this study, 316 wheat varieties (lines) were detected by high resolution melting curve (HRM) analysis and Hap-6A-P1/P2 molecular marker for TaGW2-6A allelic variations and their correlations with kernel length, kernel width, and thousand-kernel weight were analyzed. Furthermore, the TaGW2-6Aallelic variations were traced in the pedigree of the famous large-kernel variety Zhoumai 22. In the 977 bp position, 61 and 255 lines were detected with and without the T-base insertion, which were designated 977T and 977-genotypes, respectively. In the 977T genotypes, 29 lines were Hap-6A-A (TA) haplotype and 32 lines were Hap-6A-G (TG) haplotype. In the 977-genotypes, 160 lines were Hap-6A-A (-A) haplotype and 95 lines were Hap-6A-G (-G) haplotype. Significant difference was found in kernel length ( P < 0.05), kernel width ( P < 0.001) and thousand-kernel weight ( P < 0.001) between 977T and 977-genotypes. Similarly, significant difference was also found in kernel length ( P < 0.05), kernel width ( P < 0.05) and thousand-kernel weight ( P < 0.001) between Hap-6A-A and Hap-6A-G haplotypes. The allelic variation in TaGW2-6A encoding region and the promoter region jointly contributed to kernel size, and the TA haplotype was superior to -A, TG, and -G haplotypes in increasing kernel width and weight. According to pedigree analysis, the Zhoumai 22 inherited the TA haplotype from the parent Huixianhong, not from the popular parent Zhou 8425B. This haplotype is inheritable stably but tends to be lost in the process of wheat breeding. The results of this study provide not only a high-throughput molecular technique to detect wheat TA haplotype but also breeding materials in marker-assisted selection of wheat.
Keyword:Grain traits of wheat; TaGW2; Allelic variation; High resolution melting curve (HRM); Marker-assisted selection Show Figures Show Figures
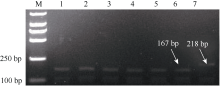
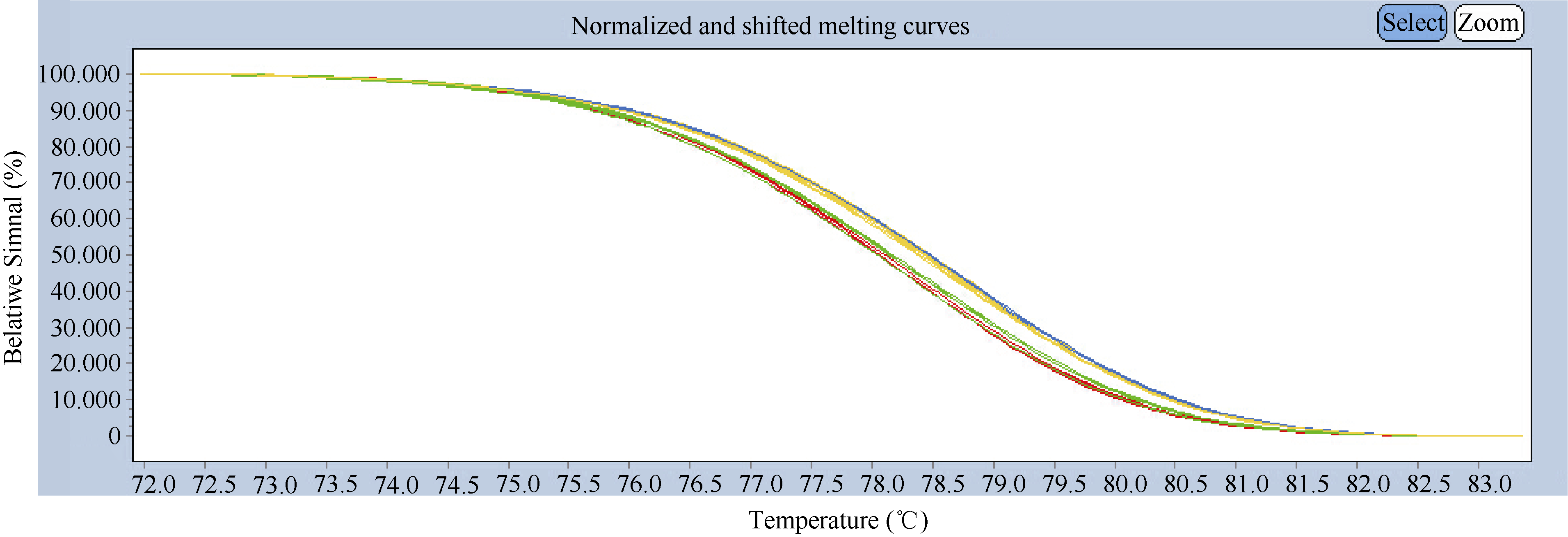
图2 977T和977-基因型的高分辨率熔解曲线分析红色、蓝色、绿色、黄色曲线分别表示兰考大粒、中国春、977T基因型和977-基因型熔解曲线。Fig. 2 High-resolution melting curve analysis of 977T and 977-genotypesThe melting curves of Lankaodali, Chinese Spring, 977T genotype, and 977-genotype are in red, blue, green, and yellow, respectively.
表1 部分重要品种名称及基因型表型数据 Table 1 Part of important varieties and their genotype-phenotype data
品种 Variety
单倍型 Haplotype
粒长 KL (mm)
粒宽 KW (mm)
粒重 TKW (g)
来源 Origin
育成年份 Year released
兰考大粒 Lankaodali
TA
8.05
3.62
51.25
中国河南 Henan, China
—
西农6028 Xinong 6028
TA
6.50
3.46
52.00
中国陕西 Shaanxi, China
1948
豫麦49 Yumai 49
TA
6.74
3.52
48.94
中国河南 Henan, China
1998
水原86 Suweon 86
TA
6.76
3.58
48.07
韩国 Public of Korea
—
辉县红 Huixianhong
TA
6.53
3.63
42.15
中国河南 Henan, China
—
陕213 Shaan 213
TA
7.30
3.22
40.98
中国陕西 Shaanxi, China
1986
加麦5号Jiamai 5
TA
6.41
3.54
48.33
加拿大 Canada
—
周麦20 Zhoumai 20
TA
6.61
3.63
56.29
中国河南 Henan, China
2003
绵7198-8 Mian 7198-8
TA
6.64
3.50
49.66
中国四川 Sichuan, China
—
西农1376 Xinong 1376
TA
6.84
3.44
40.61
中国陕西 Shaanxi, China
1988
西农2208 Xinong 2208
TA
7.57
3.16
40.87
中国陕西 Shaanxi, China
1997
周8425B Zhou 8425B
TA
7.45
3.76
52.68
中国河南 Henan, China
1988
周麦22 Zhoumai 22
TA
6.54
3.46
51.15
中国河南 Henan, China
2007
蚂蚱麦 Mazhamai
TA
6.51
3.72
36.83
中国陕西 Shaanxi, China
1930
豫麦2号 Yumai 2
TA
6.12
3.23
37.31
中国河南 Henan, China
1977
晋麦2418 Jinmai 2418
TA
6.69
3.07
43.87
中国福建 Fujian, China
—
百农3217 Bainong 3217
TA
6.70
3.02
35.16
中国河南 Henan, China
1981
冀麦2号 Jimai 2
TA
7.00
3.28
46.73
中国河北 Hebei, China
—
陕农7859 Shaannong 7859
TA
7.57
3.31
47.58
中国陕西 Shaanxi, China
1984
康定小麦 Kangding Xiaomai
TA
6.21
3.18
38.45
中国四川 Sichuan, China
—
白蚂蚱 Baimazha
TA
5.47
2.93
31.95
中国宁夏 Ningxia, China
—
青春28 Qingchun 28
TA
6.98
3.46
49.29
中国青海 Qinghai, China
1970
毕麦26 Bimai 26
TA
7.13
3.27
40.62
中国贵州 Guizhou, China
1979
去麦34 Qumai 34
TA
6.59
3.13
44.70
中国云南 Yunnan, China
—
同家坝小麦 Tongjiaba Xiaomai
TA
6.21
2.99
31.25
中国四川 Sichuan, China
—
红花麦 Honghuamai
TA
6.31
3.23
34.98
中国四川 Sichuan, China
—
成都光头 Chengduguangtou
TA
6.34
2.98
37.30
中国四川 Sichuan, China
1952
酱麦 Jiangmai
TA
6.50
3.26
36.81
中国贵州 Guizhou, China
—
汉中白 Hanzhongbai
TA
6.21
2.93
32.30
中国陕西 Shaanxi, China
—
陕253 Shaan 253
TG
7.43
3.70
42.95
中国陕西 Shaanxi, China
1994
西农2611 Xinong 2611
TG
7.21
3.35
43.76
中国陕西 Shaanxi, China
1991
武农886 Wunong 886
TG
6.38
3.33
51.75
中国陕西 Shaanxi, China
2009
邯6172 Han 6172
TG
6.69
3.25
47.77
中国河北 Hebei, China
1995
农林10号 Norin 10
TG
7.47
3.73
49.54
日本 Japan
—
临旱957 Linhan 957
TG
7.37
3.60
51.65
中国山西 Shanxi, China
—
藁优9618 Gaoyou 9618
TG
6.74
3.10
36.27
中国河北 Hebei, China
2005
扬麦5号 Yangmai 5
TG
6.94
3.00
36.69
中国江苏 Jiangsu, China
1991
西农9814 Xinong 9814
TG
6.65
3.41
52.70
中国陕西 Shaanxi, China
—
胜利麦 Triumph
TG
6.21
2.77
37.83
美国 United States
—
中农28 Villa Glori
TG
6.64
2.97
33.10
意大利 Italy
—
高原506 Gaoyuan 506
TG
6.93
3.34
46.05
中国青海 Qinghai, China
1971
宁春4号 Ningchun 4
-A
7.12
3.46
40.79
中国宁夏 Ningxia, China
1981
中麦895 Zhongmai 895
-A
7.17
3.11
47.87
中国河北 Hebei, China
2012
天麦989 Tianmai 989
-A
6.92
3.31
44.23
中国陕西 Shaanxi, China
2012
周麦12 Zhoumai 12
-A
5.99
2.99
34.50
中国河南 Henan, China
1999
周麦13 Zhoumai 13
-A
6.39
3.26
47.93
中国河南 Henan, China
—
周麦9号 Zhoumai 9
-A
5.53
2.90
38.93
中国河南 Henan, China
1980
温394 Wen 394
-A
6.96
3.15
32.40
中国河南 Henan, China
—
碧蚂6号 Bima 6
-A
5.24
3.14
36.32
中国陕西 Shaanxi, China
1948
矮丰3号 Aifeng 3
-A
6.45
3.03
33.70
中国陕西 Shaanxi, China
1970
咸农39 Xiannong 39
-A
7.01
3.09
37.44
中国陕西 Shaanxi, China
1964
小偃6号 Xiaoyan 6
-A
6.63
3.38
43.09
中国陕西 Shaanxi, China
1981
小偃22 Xiaoyan 22
-A
6.97
3.28
38.86
中国陕西 Shaanxi, China
2003
郑麦9023 Zhengmai 9023
-A
7.12
3.15
46.36
中国河南 Henan, China
2009
中麦349 Zhongmai 349
-A
6.62
2.79
42.85
中国河北 Hebei, China
2009
新麦9号 Xinmai 9
-A
6.49
3.26
42.99
中国河南 Henan, China
1997
存麦1号 Cunmai 1
-A
6.76
3.38
44.25
中国河南 Henan, China
2011
浚麦55 Xunmai 55
-A
6.27
3.06
46.70
中国河南 Henan, China
—
淮麦20 Huaimai 20
-A
7.27
3.46
44.07
中国江苏 Jiangsu, China
2003
丰产3号 Fengchan 3
-G
6.34
3.13
36.86
中国陕西 Shaanxi, China
1964
鲁麦1号 Lumai 1
-G
6.19
3.21
38.13
中国山东 Shandong, China
1975
Data are the means over two years (2012-2013 and 2013-2014 wheat seasons). “ — ” indicates released year unknown. 数据为2012-2013和2013-2014两年度的平均值。— 表示育成时间未知。
表1 部分重要品种名称及基因型表型数据 Table 1 Part of important varieties and their genotype-phenotype data
表2TaGW2-6A基因等位变异的表型关联分析 Table 2 Correlation analysis of TaGW2-6A polymorphism and phenotype
基因型/单倍型 Genotype/haplotype
粒长 Kernel length (mm)
粒宽 Kernel width (mm)
千粒重 1000-kernel weight (g)
2012-2013
977T
6.67± 0.07 aA
3.30± 0.03 aA
42.40± 0.82 aA
977-
6.41± 0.03 bA
3.13± 0.02 bB
38.07± 0.40 bB
Hap-6A-A
6.52± 0.04 aA
3.19± 0.02 aA
40.46± 0.47 aA
Hap-6A-G
6.39± 0.05 bA
3.11± 0.02 bA
36.60± 0.57 bB
TA
6.73± 0.10 aA
3.36± 0.05 aA
43.43± 1.19 aA
-A
6.48± 0.04 bA
3.17± 0.02 bB
39.96± 0.51 bB
TG
6.62± 0.10 abA
3.25± 0.05 abAB
41.46± 1.13 abAB
-G
6.31± 0.06 cC
3.07± 0.03 cB
34.96± 0.66 cC
2013-2014
977T
6.61± 0.07 aA
3.26± 0.03 aA
41.66± 0.80 aA
977-
6.35± 0.03 bA
3.10± 0.02 bB
37.72± 0.39 bB
Hap-6A-A
6.45± 0.04 aA
3.16± 0.02 aA
39.95± 0.45 aA
Hap-6A-G
6.32± 0.05 bA
3.09± 0.02 bA
36.29± 0.55 bB
TA
6.68± 0.10 aA
3.29± 0.05 aA
42.64± 1.16 aA
-A
6.41± 0.04 bA
3.14± 0.02 bA
39.46± 0.49 bA
TG
6.54± 0.10 abA
3.23± 0.05 abA
40.77± 1.10 abA
-G
6.24± 0.06 cB
3.04± 0.03 cB
34.78± 0.64 cB
In each growing year, values followed by different capitals and lowercases are significantly different between TaGW2-6A genotypes or among TaGW2-6A haplotypes at P < 0.001 and P < 0.05, respectively. 同一年度中, 不同大写和小写字母分别表示TaGW2-6A位点2种基因型之间或启动子区4种单倍型之间在0.001和0.05概率水平下差异显著。
表2TaGW2-6A基因等位变异的表型关联分析 Table 2 Correlation analysis of TaGW2-6A polymorphism and phenotype
Fox JL. Whatever happened to GM wheat?Nat Biotechnol, 2009, 27: 974-976[本文引用:1]
[2]
庄巧生. 中国小麦品种改良及系谱分析. 北京: 中国农业出版社, 2003. pp 497-518Zhuang QS. Analysis of Chinese Wheat Impronement and Pedigree. Beijing: China Agriculture Press, 2003. pp 497-518(in Chinese)[本文引用:1]
[3]
许为刚, 胡琳, 吴兆苏, 盖钧镒. 关中地区小麦品种产量与产量结构遗传改良的研究. 作物学报, 2000, 26: 352-358Xu WG, HuL, Wu ZS, Gai JY. Studies on genetic improvement of yield and yield components of wheat cultivars in Mid-Shaanxi area. Acta Agron Sin, 2000, 26: 352-358 (in Chinese with English abstract)[本文引用:1]
[4]
胡延吉, 赵檀方. 小麦高产育种中粒重作用的研究. 作物学报, 1995, 21: 671-678Hu YJ, Zhao TF. Studies on the effect of grain weight in breeding of high-yield wheat. Acta Agron Sin, 1995, 21: 671-678 (in Chinese with English abstract)[本文引用:1]
[5]
WuW, Li CJ, Ma BL, ShahF, LiuY, Liao YC. Genetic progress in wheat yield and associated traits in China since 1945 and future prospects. Euphytica, 2013, 196: 155-168[本文引用:1]
[6]
Song XJ, HuangW, ShiM, Zhu MZ, Lin HX. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet, 2007, 39: 623-630[本文引用:1]
[7]
姜颖, 曹言勇, 路运才, 唐保军, 王利锋, 李会勇. 拟南芥中RING型E3泛素连接酶基因AtGW2的克隆和功能分析. 植物遗传资源学报, 2011, 12: 448-454JiangY, Cao YY, Lu YC, Tang BJ, Wang LF, Li HY. Cloning and functional analysis of Arabidopsis thaliana AtGW2, a RING-type E3 ubiquitin ligase protein. J Plant Genet Resours, 2011, 12: 448-454 (in Chinese with English abstract)[本文引用:1]
[8]
LiQ, LiL, Yang XH, Warburton ML, Bai GH, Dai JR, Li JS, Yan JB. Relationship, evolutionary fate and function of two maize co-orthologs of rice GW2 associated with kernel size and weight. BMC Plant Biol, 2010, 10: 1-15[本文引用:1]
[9]
Hong YT, Chen LF, Du LP, Su ZQ, Wang JF, Ye XG, LiQ, Zhang ZY. Transcript suppression of TaGW2 increased grain width and weight in bread wheat. Funct Integr Genomics, 2014, 14: 341-349[本文引用:1]
[10]
Yang ZB, Bai ZY, Li XL, WangP, Wu QX, YangL, Li LQ, Li XJ. SNP identification and allelic-specific PCR markers development for TaGW2, a gene linked to wheat kernel weight. Theor Appl Genet, 2012, 125: 1057-1068[本文引用:3]
[11]
Su ZQ, Hao CY, Wang LF, Dong YC, Zhang XY. Identification and development of a functional marker of TaGW2 associated with grain weight in bread wheat (Triticum aestivum L. ). Theor Appl Genet, 2011, 122: 211-223[本文引用:2]
[12]
Gawel NJ, Jarret RL. A modified CTAB DNA extraction procedure for Musa and Ipomoea. Plant Mol Biol Rep, 1991, 9: 262-266[本文引用:2]
[13]
LiuJ, Huang SM, Sun MY, Liu SY, Liu YM, Wang WX, Zhang XR, Wang HZ, HuaW. An improved allele-specific PCR primer design method for SNP marker analysis and its application. Plant Methods, 2012, 8: 1-9[本文引用:1]
[14]
Xu YB, Crouch JH. Marker-assisted selection in plant breeding: from publications to practice. Crop Sci, 2008, 48: 391-407[本文引用:1]
[15]
Gundry CN, Vand ersteen JG, Reed GH, Pryor RJ, ChenJ, Wittwer CT. Amplicon melting analysis with labeled primers: a closed-tube method for differentiating homozygotes and heterozygotes. Mol Diagnostics and Genet, 2003, 49: 396-406[本文引用:1]
GanopoulosI, BosmaliI, MadesisP, TsaftarisA. Microsatellite genotyping with HRM (high resolution melting) analysis for identification of the PGI common bean variety Plake Megalosperma Prespon. Eur Food Res Technol, 2012, 234: 501-508[本文引用:1]
[18]
Jeong HJ, Jo YD, Park SW, Kang BC. Identification of Capsicum species using SNP markers based on high resolution melting analysis. Genome, 2010, 53: 1029-1040[本文引用:1]
[19]
Han YH, Khu DM, Monteros MJ. High-resolution melting analysis for SNP genotyping and mapping in tetraploid alfalfa (Medicago sativa L. ). Mol Breed, 2011, 29: 489-501[本文引用:1]
[20]
Hofinger BJ, Jing HC, Hammond-KosackK E, KanyukaK. High-resolution melting analysis of cDNA-derived PCR amplicons for rapid and cost-effective identification of novel alleles in barley. Theor Appl Genet, 2009, 119: 851-865[本文引用:1]
 , 高欣, 李立群, 李扬, 王中华, 李学军
, 高欣, 李立群, 李扬, 王中华, 李学军




{kind=link}
{kind=link}
{kind=link}
{kind=link}