关键词:水稻; 斑点叶突变体; 过氧化氢; 光合色素; 基因定位 Characterization and Gene Mapping of a Spotted-leaf Mutant spl21 in Rice ( Oryza sativaL.) SONG Li-Xin1,2, HUANG Qi-Na1, FENG Bao-Hua1, SHI Yong-Feng1, ZHANG Xiao-Bo1, XU Xia1, WANG Hui-Mei1, LI Xiao-Hong1, ZHAO Bao-Hua2,*, WU Jian-Li1,* 1 State Key Laboratory of Rice Biology / Chinese National Center for Rice Improvement / China National Rice Research Institute, Hangzhou 310006, China
2 College of Life Sciences, Hebei Normal University, Shijiazhuang 050024, China
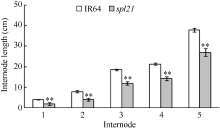
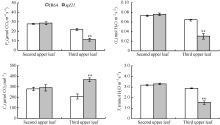
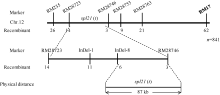
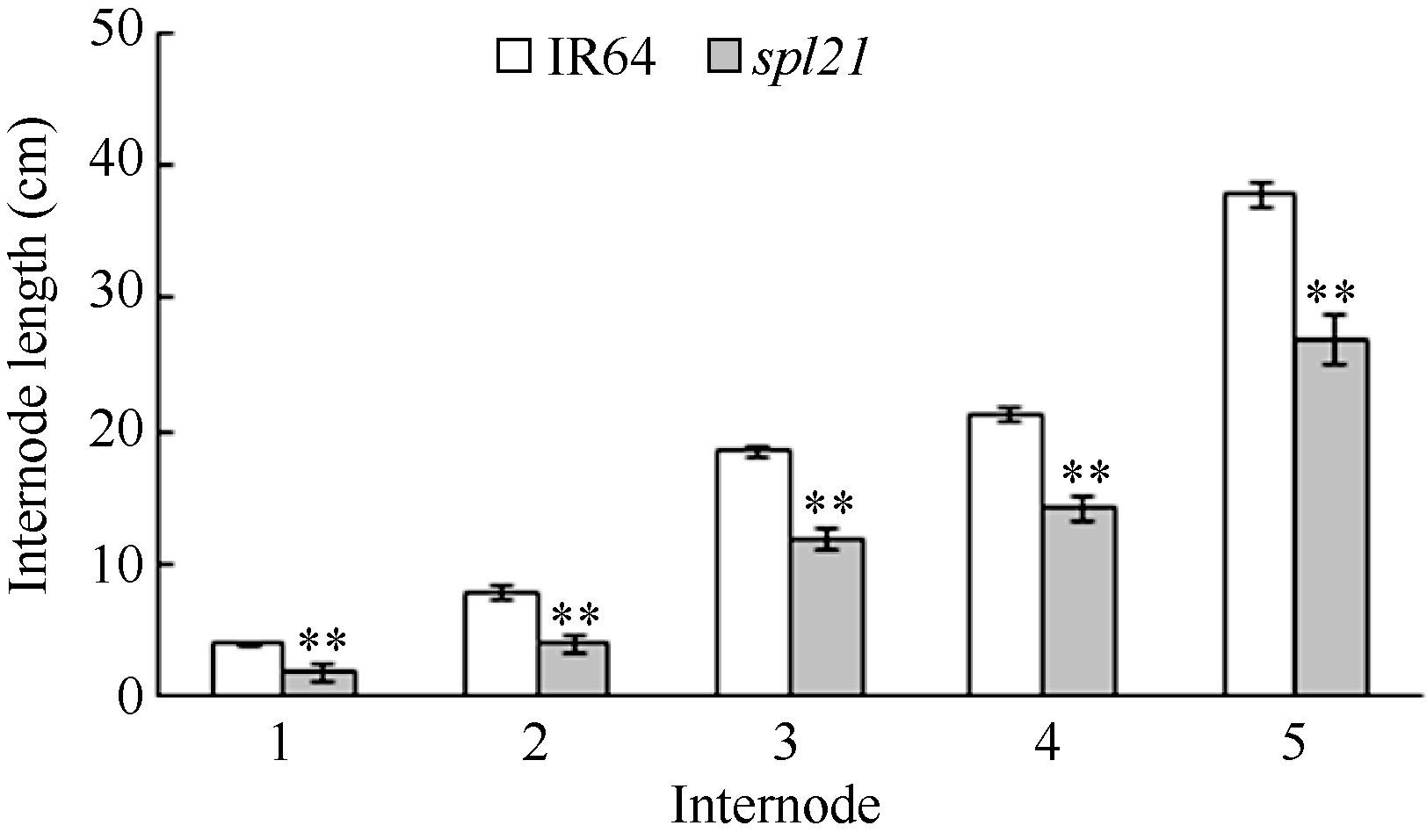
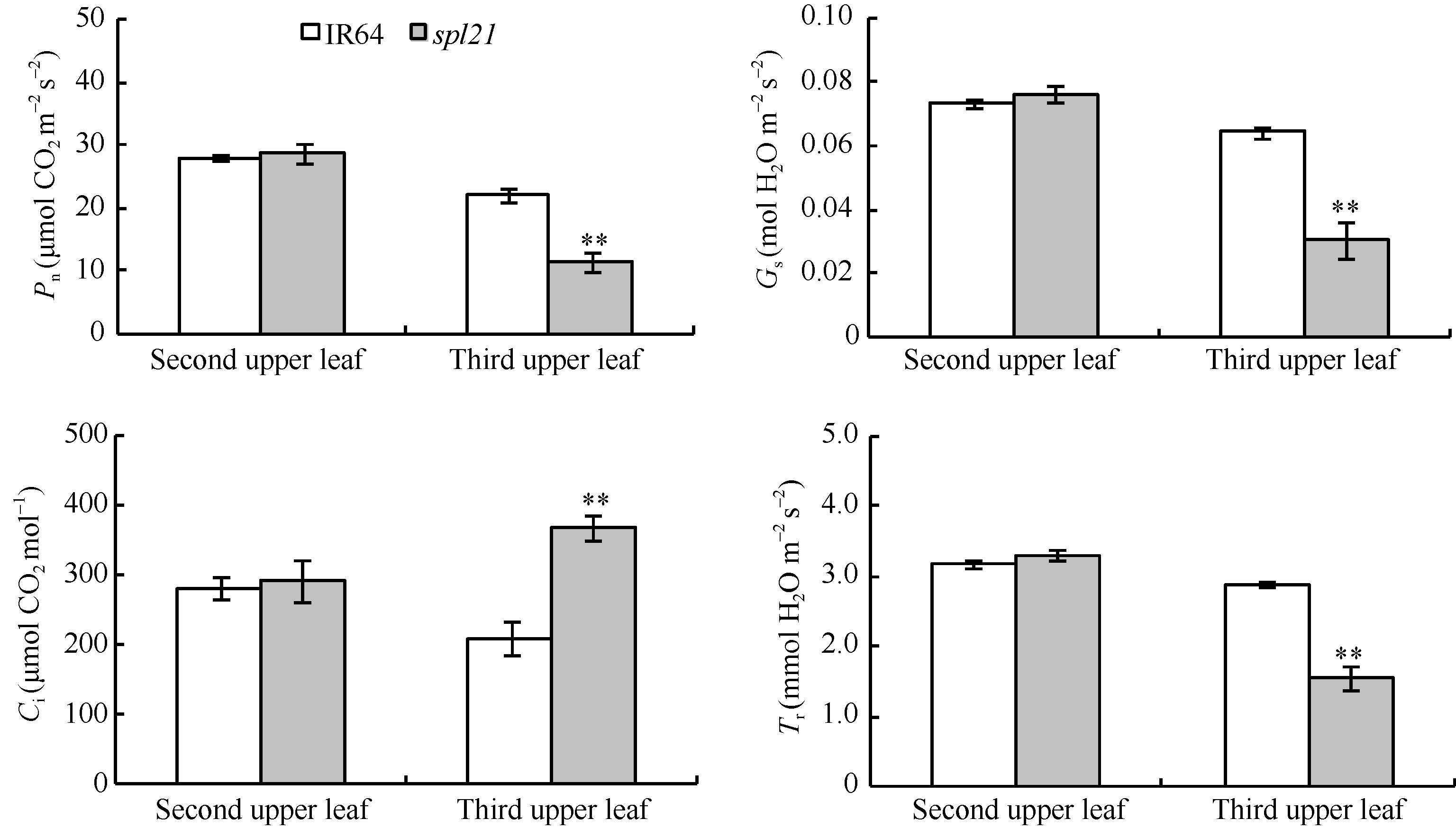
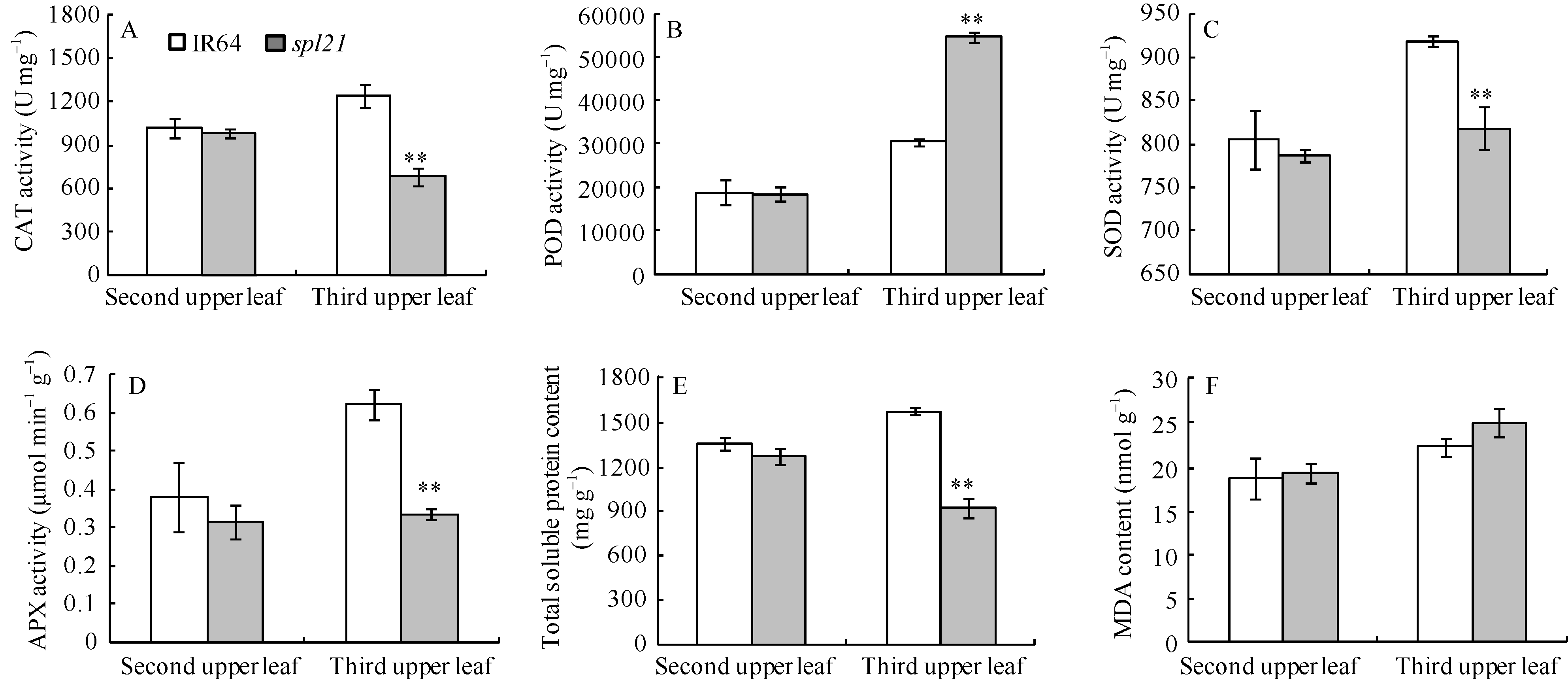
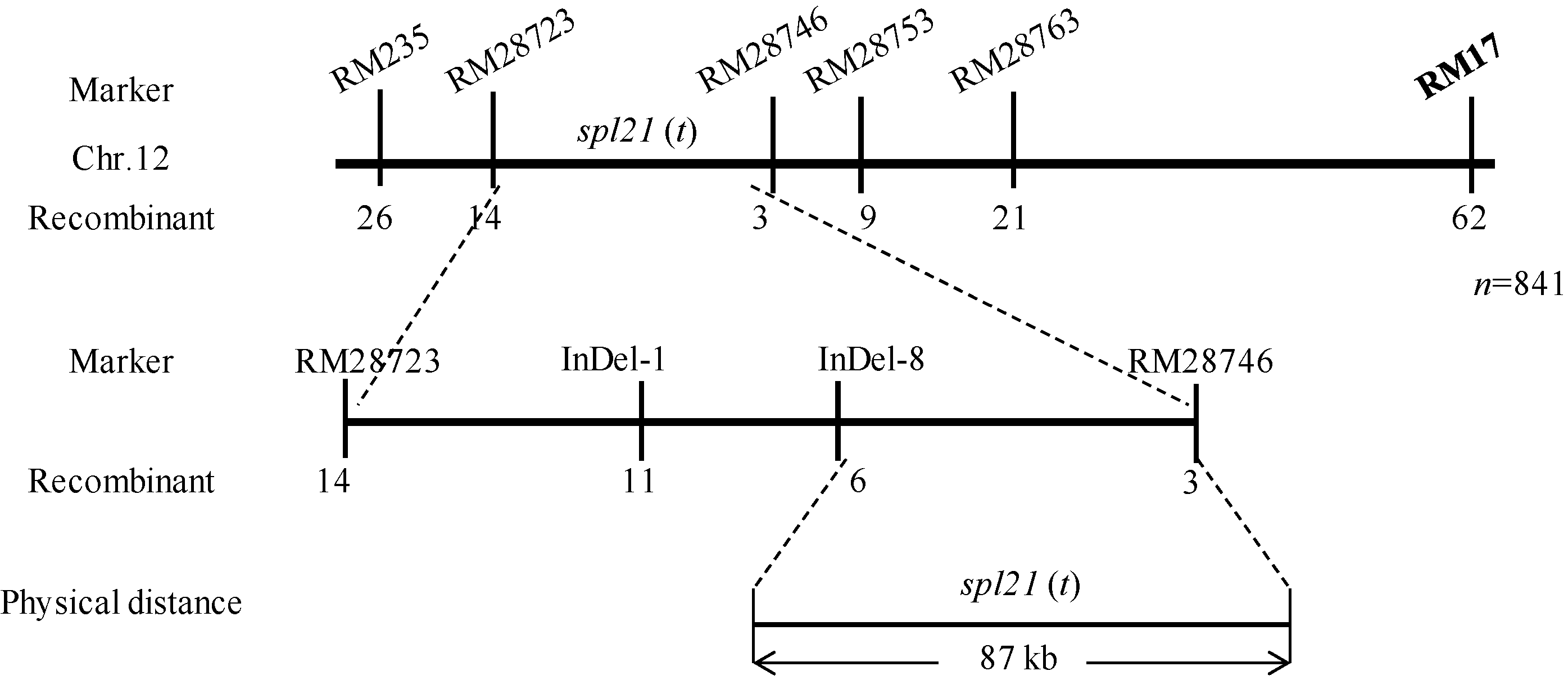
AbstractThe rice spotted-leaf 21 mutant ( spl21) was isolated from a diepoxybutane-induced IR64 mutant bank. Under field conditions, the red-brown spots appeared on the leaves of mutant seedlings in two weeks after sowing. Subsequently, a portion of spots merged and the leaf tips became yellowish, wilted and spread downwards along both edges of the leaf blade leading to the death of the whole leaf blade when the symptom was severe. Accumulation of H2O2 was detected in and around the spots. Major agronomic traits including plant height, length of panicle, number of panicles, number of filled grains, seed setting-rate, and 1000-grain weight were markedly affected in the mutant. The contents of chlorophyll a, b, carotenoid and photosynthetic parameters were significantly reduced in the mutant as compared with the wild type. Furthermore, the activities of CAT, SOD, APX, and soluble protein contents were significantly lower than those of the wild type while the activity of POD was apparently higher than that of the wild type. The mutant trait was controlled by a single recessive nuclear gene, tentatively termed spl21( t), located on the long arm of chromosome 12. The population and data achieved in the present study would facilitate the isolation and functional analysis of spl21( t).
Keyword:Rice; Spotted-leaf mutant; Hydrogen peroxide; Photosynthetic pigment; Gene mapping Show Figures Show Figures
图1 突变体表型A: 野生型IR64幼苗; B: 突变体spl21幼苗; C: 苗期叶片 (左: IR64; 右: spl21); D: 分蘖期IR64和spl21。Fig. 1 Phenotype of the mutantA: seedling of IR64; B: seedling of the mutant; C: leaves at the seedling stage (Left: IR64; Right: spl21); D: plants of the wild type and the mutant at the tillering stage.
表1 突变体和野生型的农艺性状 Table 1 Performance of agronomic traits in the mutant and wild type
材料 Material
株高 Plant height (cm)
有效穗数 No. of panicles
穗长 Panicle length (cm)
实粒数 Filled grain/panicle
结实率 Seed-setting rate (%)
千粒重 1000-grain weight (g)
IR64
118.9± 1.5
19.3± 1.2
26.6± 0.5
1471.3± 281.5
72.5± 5.8
27.2± 0.4
spl21
89.1± 6.5* *
13.7± 2.8* *
24.1± 1.0*
370.3± 42.6* *
37.3± 2.8* *
20.2± 1.2* *
* and * * denote significant difference at the 0.05 and 0.01 probability levels, respectively. * 和* * 表示在0.05和0.01水平上差异显著。
表1 突变体和野生型的农艺性状 Table 1 Performance of agronomic traits in the mutant and wild type
表2 Table 2 表2(Table 2)
表2 苗期叶片光合色素含量 Table 2 Pigment contents at the seedling stage
材料 Material
叶绿素a Chlorophyll a (mg g-1)
叶绿素b Chlorophyll b (mg g-1)
叶绿素a/b Chlorophyll a/b
类胡萝卜素 Carotenoid (mg g-1)
IR64
1.994± 0.105 a
0.580± 0.032 a
3.438± 0.046 a
0.373± 0.016 a
spl21-NO
2.154± 0.423 a
0.629± 0.107 a
3.420± 0.183 a
0.412± 0.089 a
spl21-SPL
1.107± 0.197 b
0.352± 0.065 b
3.143± 0.086 a
0.217± 0.030 b
spl21-NO: mutant leaves without lesions; spl21-SPL: mutant leaves with lesions. Values followed by different letters are significantly difference at P≤ 0.05 on the basis of Duncan’ s test. spl21-NO: 无斑点的突变体叶片; spl21-SPL: 有红褐斑的突变体叶片。a、b表示Duncan’ s测验显著(P≤ 0.05)。
表2 苗期叶片光合色素含量 Table 2 Pigment contents at the seedling stage
图3 突变体spl21和野生型IR64叶片DAB染色结果A: IR64染色之前; B: IR64染色之后; C: spl21无斑叶片(spl21-NO)DAB染色之前; D: spl21无斑叶片(spl21-NO)DAB染色之后; E: spl21褐斑叶片(spl21-SPL)DAB染色之前; F: spl21褐斑叶片(spl21-SPL)DAB染色之后。Fig. 3 DAB staining of leaves in the mutant spl21 and wild type IR64A: IR64 before DAB staining; B: IR64 after DAB staining; C: spl21-NO before DAB staining; D: spl21-NO after DAB staining; E: spl21-SPL before DAB staining; F:spl21-SPL after DAB staining.
黄奇娜, 杨杨, 施勇烽, 陈洁, 吴建利. 水稻斑点叶变异研究进展. 中国水稻科学, 2010, 24: 108-115Huang QN, YangY, Shi YF, ChenJ, Wu JL. Recent advances in research on spotted leaf mutants of rice (Oryza sativa). Chin J Rice Sci, 2010, 24: 108-115 (in Chinese with English abstract)[本文引用:3]
[2]
Huang QN, Shi YF, YangY, Feng BH, Wei YL, ChenJ, MariettaB, LeungH, Wu JL. Characterization and genetic analysis of a light and temperature-sensitive spotted-leaf mutant in rice. J Integr Plant Biol, 2011, 53: 671-681[本文引用:3]
[3]
Feng BH, YangY, Shi YF, Shen HC, Wang HM, Huang QN, XuX, Lv XG, Wu JL. Characterization and genetic analysis of a novel rice spotted-leaf mutant HM47 with broad-spectrum resistance to Xanthomonas oryzae pv. oryzae. J Integr Plant Biol, 2013, 55: 473-483[本文引用:1]
[4]
Dietrich RA, Richberg MH, SchmidtR, DeanC, Dangl JL. A novel zinc finger protein is encoded by the arabidopsis LSD1 gene and functions as a negative regulator of plant cell death. Cell, 1997, 88: 685-694[本文引用:2]
[5]
GrayJ, Close PS, Briggs SP, Johal GS. A novel suppressor of cell death in plants encoded by the LIS1 gene of maize. Cell, 1997, 89: 25-31[本文引用:3]
[6]
BuschgesR, HollricherK, PanstrugaR, SimonsG, WolterM, FrijtersA, DaelenR, LeeT, DiergaardeP, GroenendijkJ, TopschS, VosP, SalaminiF, Schulze LP. The barley mlo gene: a novel control element of plant pathogen resistance. Cell, 1997, 88: 695-705[本文引用:1]
[7]
Badigannavar AM, Kale DM, EapenS, Murty GS. Inheritance of disease lesion mimic leaf trait in groundnut. J Hered, 2002, 93: 50-52[本文引用:1]
[8]
陈析丰, 金杨, 马伯军. 水稻类病变突变体及抗病性的研究进展. 植物病理学报, 2011, 41: 1-9Chen XF, JinY, Ma BJ. Progress on the studies of rice lesion mimics and their resistant mechanism to the pathogens. Acta Phytopathol Sin, 2011, 41: 1-9 (in Chinese with English abstract)[本文引用:2]
[9]
邱结华, 马宁, 蒋汉伟, 圣忠华, 邵高能, 唐绍清, 魏祥进, 胡培松. 水稻类病斑突变体lmm4的鉴定及其基因定位. 中国水稻科学, 2014, 28: 367-376Qiu JH, MaN, Jiang HW, Sheng ZH, Shao GN, Tang SQ, Wei XJ, Hu PS. Identification and gene mapping of a lesion mimic mutant lmm4 in rice. Chin J Rice Sci, 2014, 28: 367-376 (in Chinese with English abstract)[本文引用:1]
[10]
LiZ, Zhang YX, LiuL, LiuQ, Bi ZB, YuN, Cheng SH, Cao LY. Fine mapping of the lesion mimic and early senescence 1 (lmes1) in rice (Oryza sativa). Plant Physiol Biochem, 2014, 80: 300-307[本文引用:1]
[11]
XuX, Zhang LL, Liu BM, Ye FY, Wu YJ. Characterization and mapping of a spotted leaf mutant in rice (Oryza sativa). Genet Mol Biol, 2014, 37: 406-413[本文引用:1]
[12]
刘林, 张迎信, 李枝, 刘群恩, 余宁, 孙滨, 杨正福, 周全, 程式华, 曹立勇. 水稻类病变突变体g303的鉴定和基因定位. 中国水稻科学, 2014, 28: 465-472LiuL, Zhang YX, LiZ, Liu QN, YuN, SunB, Yang ZF, ZhouQ, Cheng SH, Cao LY. Characterization and gene mapping of a lesion mimic mutant g303 in rice. Chin J Rice Sci, 2014, 28: 465-472 (in Chinese with English abstract)[本文引用:1]
[13]
韩雪颖, 杨勇, 余初浪, 张文浩, 叶胜海, 陈斌, 程晨, 程晔, 严成其, 陈剑平. 一个抗病性增强的水稻类病变突变体的蛋白质组学研究. 中国水稻科学, 2014, 28: 559-569Han XY, YangY, Yu CL, Zhang WH, Ye SH, ChenB, ChengC, ChengY, Yan CQ, Chen JP. A proteomic study on a disease-resistance-enhanced rice lesion mimic mutant. Chin J Rice Sci, 2014, 28: 559-569 (in Chinese with English abstract)[本文引用:2]
[14]
YamanouchiU, YanoM, LinH, AshikariM, YamadaK. A rice spotted leaf gene, Spl7, encodes a heat stress transcription factor protein. Proc Natl Acad Sci USA, 2002, 99: 7530-7535[本文引用:2]
[15]
Zeng LR, QuS, BordeosA, YangC, BaraoidanM, YanH, XieQ, Nahm BH, LeungH, Wang GL. Spotted leaf 11, a negative regulator of plant cell death and defense, encodes a U-box/ armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell, 2004, 16: 2795-2808[本文引用:3]
[16]
MoriM, TomitaC, SugimotoK, HasegawaM, HayashiN, Dubouzet JG, OchiaiH, SekimotoH, HirochikaH, KikuchiS. Isolation and molecular characterization of a spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol Biol, 2007, 63: 847-860[本文引用:1]
[17]
QiaoY, JiangW, LeeJ, ParkB, Choi MS, PiaoR, Woo MO, Roh JH, HanL, Paek NC, Seo HS, Koh HJ. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit l1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol, 2010, 185: 258-274[本文引用:2]
[18]
Wang LY, Pei ZY, Tian YC, He CZ. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation. Mol Plant Microbe Interact, 2005, 18: 375-384[本文引用:2]
[19]
Kim JA, ChoK, SinghR, Jung YH, Jeong SH, Kim SH, Lee JE, Cho YS, Agrawal GK, RakwalR, TamogamiS, KerstenB, Jeon JS, AnG, Jwa NS. Rice OsACDR1 (Oryza sativa accelerated cell death and resistance 1) is a potential positive regulator of fungal disease resistance. Mol Cells, 2009: 431-439[本文引用:1]
[20]
Yuan YX, Zhong SH, LiQ, Zhu ZR, Lou YL, Wang LY, Wang JJ, Wang MY, Li QL, Yang DL, He ZH. Functional analysis of rice NPR1-like genes reveals that OsNPR1/NHI is the rice orthologue conferring disease resistance with enhanced herbivore susceptibility. Plant Biotechnol J, 2007, 5: 313-324[本文引用:1]
Sun CH, Liu LC, Tang JY, LinA, Zhang FT, FangJ, Zhang GF, Chu CC. Rlin1, encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice. J Genet Genomics, 2011, 38: 29-37[本文引用:1]
[23]
Tang JY, Zhu XD, Wang YQ, Liu LC, XuB, LiF, FangJ, Chu CC. Semi-dominant mutations in the CC-NB-LRR-type R gene, NLS1, lead to constitutive activation of defense responses in rice. Plant J, 2011, 66: 996-1007[本文引用:1]
[24]
Liu XQ, LiF, Tang JY, Wang WH, Zhang FX, Wang GD, Chu JF, Yan CY, Wang TQ, Chu CC, Li CY. Activation of the jasmonic acid pathway by depletion of the hydroperoxide lyase OsHPL3 reveals crosstalk between the HPL and AOS branches of the oxylipin pathway in rice. Plos One, 2012, 7: 1-14[本文引用:1]
[25]
Chen XF, HaoL, Pan JW, Zheng XX, Jiang GH, JinY, Gu ZM, QianQ, Zhai WX, Ma BJ. SPL5, a cell death and defense-related gene, encodes a putativesplicing factor 3b subunit 3 (SF3b3) in rice. Mol Breed, 2012, 30: 939-949[本文引用:1]
[26]
Undan JR, TamiruM, AbeA, YoshidaK, KosugiS, TakagiH, YoshidaK, KanzakiH, SaitohH, FekihR, SharmaS, UndanJ, YanoM, TerauchiR. Mutation in OsLMS, a gene encoding a protein with two double-strand ed RNA binding motifs, causes lesion mimic phenotype and early senescence in rice (Oryza sativa L. ). Genes Genet Syst, 2012, 87: 169-179[本文引用:1]
[27]
LinA, WangY, TangJ, XueP, LiC, LiuL, HuB, YangF, Loake GJ, ChuC. Nitric oxide and protein s-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice. Plant Physiol, 2012, 158: 451-464[本文引用:1]
[28]
SakurabaY, Rahman ML, Cho SH, Kim YS, Koh HJ, Yoo SC, Paek NC. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J, 2013, 74: 122-133[本文引用:1]
[29]
FujiwaraT, MaisonneuveS, IsshikiM, MizutaniM, ChenL, Wong HL, KawasakiT, ShimamotoK. Sekiguchi lesion gene encodes a cytochrome P450 monooxygenase that catalyzes conversion of tryptamine to serotonin in rice. J Biol Chem, 2010, 285: 11308-11313[本文引用:2]
[30]
Jiao BB, Wang JJ, Zhu XD, Zeng LJ, LiQ, He ZH. A novel protein RLS1 with NB-ARM domainsis involved in chloroplast degradation during leaf senescence in rice. Mol Plant, 2012, 5: 205-217[本文引用:1]
[31]
FekihR, TamiruM, KanzakiH, AbeA, YoshidaK, KanzakiE, SaitohH, TakagiH, NatsumeS, Undan JR, UndanJ, TerauchiR. The rice (Oryza sativa L. ) LESION MIMIC RESEMBLING, which encodes an AAA-type ATPase, is implicated in defense response. Mol Genet Genomics, 2015, 290: 611-622[本文引用:1]
[32]
BalagueC, LinB, AlconC, FlottesG, MalmstromS, KohlerC, NeuhausG, PelletierG, GaymardF, RobyD. HLM1, an essential signaling component in the hypersensitive response, is a member of the cyclic nucleotide-gated channel ion channel family. Plant Cell, 2003, 15: 365-379[本文引用:1]
[33]
HuG, YalpaniN, Briggs SP, Johal GS. A porphyrin pathway impairment is responsible for the phenotype of a dominant disease lesion mimic mutant of maize. Plant Cell, 1998, 10: 1095-1105[本文引用:1]
[34]
BrodersenP, Malinovsky FG, HematyK, Newman MA, MundyJ. The role of salicylic acid in the induction of cell death in Arabidopsis acd11. Plant Physiol, 2005, 138: 1037-1045[本文引用:1]
[35]
Wu CJ, BordeosA, Madamba MS, BaraoidanM, RamosM, Wang GL, Leach JE, LeungH. Rice lesion mimic mutants with enhanced resistance to diseases. Mol Genet Genomics, 2008, 276: 605-619[本文引用:1]
[36]
Arnon DI. Copper enzymes in isolated chloroplasts polyphenoloxidase in Beta vulgaris. Plant Physiol, 1949, 24: 1-15[本文引用:1]
[37]
Wellburn AR. The spectral determination of chlorophyll a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. Plant Physiol, 1994, 144: 307-313[本文引用:1]
[38]
Thordal-ChristansenH, Zhang ZG, Wei YD, Collinge DB. Subcellular localization of H2O2 in plants H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J, 1997, 11: 1187-1194[本文引用:1]
[39]
赵世杰, 史国安, 董新纯. 植物生理学实验指导. 北京: 中国农业科学技术出版社, 2002. pp 134-143Zhao SJ, Shi GA, Dong XC. Plant Physiology Experiment Instruction. Beijing: China Agricultural Science and Technology Press, 2002. pp 134-143(in Chinese)[本文引用:2]
[40]
卢扬江, 郑康乐. 提取水稻DNA的一种简易方法. 中国水稻科学, 1992, 6: 47-48Lu YJ, Zheng KL. A simple method for isolation of rice mitochondrial DNA. Chin J Rice Sci, 1992, 6: 47-48 (in Chinese with English abstract)[本文引用:1]
[41]
李梦钗, 冯薇, 葛艳蕊. 臭氧处理对草莓果实PPO和POD活性的影响. 经济林研究, 2012, 30(3): 84-86Li MC, FengW, Ge YR. Effects of ozone treatment on PPO and POD activities in strawberry fruit. Nonwood For Res, 2012, 30(3): 84-86 (in Chinese with English abstract)[本文引用:1]
[42]
李秀兰, 王平荣, 曲志才, 孙小秋, 王兵, 邓晓建. 水稻类病变突变体C23的遗传分析与基因的精细定位. 中国农业科学, 2010, 43: 3691-3697Li XL, Wang PR, Qu ZC, Sun XQ, WangB, Deng XJ. Genetic analysis and fine mapping of a lesion mimic mutant C23 in rice. Sci Agric Sin, 2010, 43: 3691-3697 (in Chinese with English abstract)[本文引用:1]
[43]
杨绍华, 刘华清, 王锋. 水稻斑点叶突变体W1764的遗传分析及初步定位. 福建农业学报, 2011, 26: 519-522Yang SH, Liu HQ, WangF. Genetic analysis and gene mapping of a spotted leaf mutant W1764 in rice. Fujian J Agric Sci, 2011, 26: 519-522 (in Chinese with English abstract)[本文引用:1]
[44]
吴超, 付亚萍, 胡国成, 斯华敏, 刘旭日, 孙宗修, 程式华, 刘文真. 一个水稻类病变黄叶突变体的鉴定和精细定位. 中国水稻科学, 2011, 25: 256-260WuC, Fu YP, Hu GC, Si HM, Liu XR, Sun ZX, Cheng SH, Liu WZ. Identification and fine mapping of a spotted and yellow leaf mutant in rice. Chin J Rice Sci, 2011, 25: 256-260 (in Chinese with English abstract)[本文引用:1]
[45]
陈萍萍, 叶胜海, 赵宁春, 陆艳婷, 刘合芹, 杨玲, 金庆生, 张小明. 浙粳22类病斑突变体spl(t)特征及其基因定位. 核农学报, 2010, 24: 1-6Chen PP, Ye SH, Zhao NC, Lu YT, Liu HQ, YangL, Jin QS, Zhang XM. Characteristics and genetic mapping of a lesion mimic mutant spl(t) in japonica rice variety Zhejing 22. J Nucl Agric Sci, 2010, 24: 1-6 (in Chinese with English abstract)[本文引用:1]
[46]
代高猛, 朱小燕, 李云峰, 凌英华, 赵芳明, 杨正林, 何光华. 水稻类病斑突变体spl31的遗传分析与基因定位. 作物学报, 2013, 39: 1223-1230Dai GM, Zhu XY, Li YF, Ling YH, Zhao FM, Yang ZL, He GH. Genetic analysis and fine mapping of a lesion mimic mutant spl31 in rice. Acta Agron Sin, 2013, 39: 1223-1230 (in Chinese with English abstract)[本文引用:1]
[47]
龙继凤, 潘英华, 秦学毅, 罗兴录, 朱汝财. 水稻类病变坏死突变体的形态观察及基因初步分析. 广西农业科学, 2009, 40: 614-617Long JF, Pan YH, Qin XY, Luo XL, Zhu RC. Morphological observation and gene analysis of lesion mimic mutant of rice (Oryza sativa L. ). Guangxi Agric Sci, 2009, 40: 614-617 (in Chinese with English abstract)[本文引用:1]
 , 黄奇娜
, 黄奇娜





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}