关键词:水稻; 黄绿叶突变体; 遗传分析; 精细定位 Genetic Analysis of a New Yellow-green Leaf Mutant and Fine-mapping of Mutant Gene in Rice HE Ni-Qing, LIU Zhou, ZHANG Long, BAI Su-Yang, TIAN Yun-Lu, JIANG Ling*, WAN Jian-Min State Key Laboratory of Crop Genetics and Germplasm Enhancement / Research Center of Jiangsu Plant Gene Engineering / Ministry of Agriculture Key Laboratory of Biology / Genetics and Breeding of Japonica Rice in Mid-lower Yangtze River / The Yangtze River Valley Hybrid Rice Collaboration Innovation Center, Nanjing Agricultural University, Nanjing 210095, China
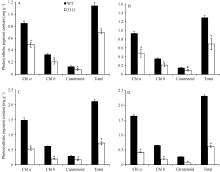
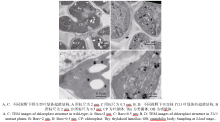
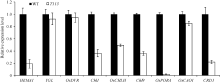
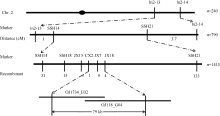
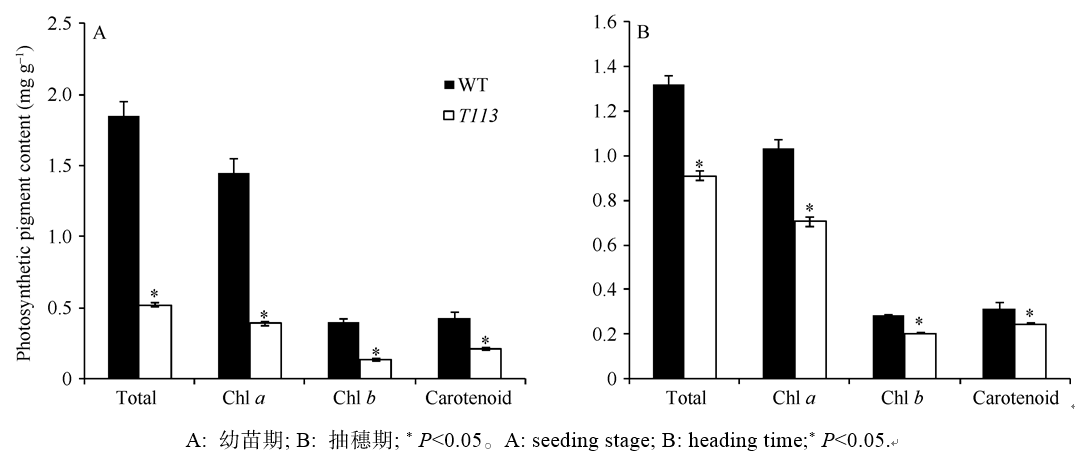
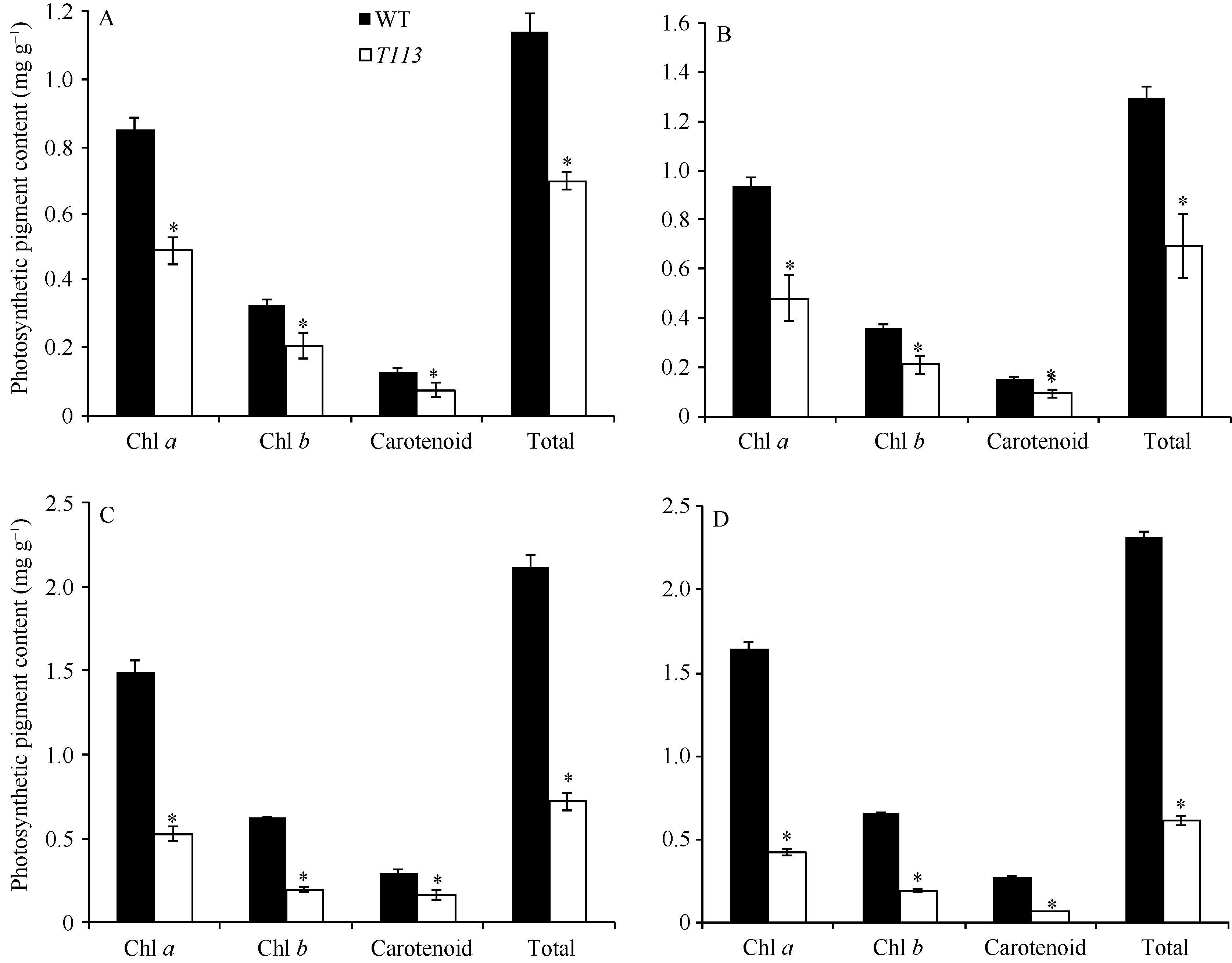
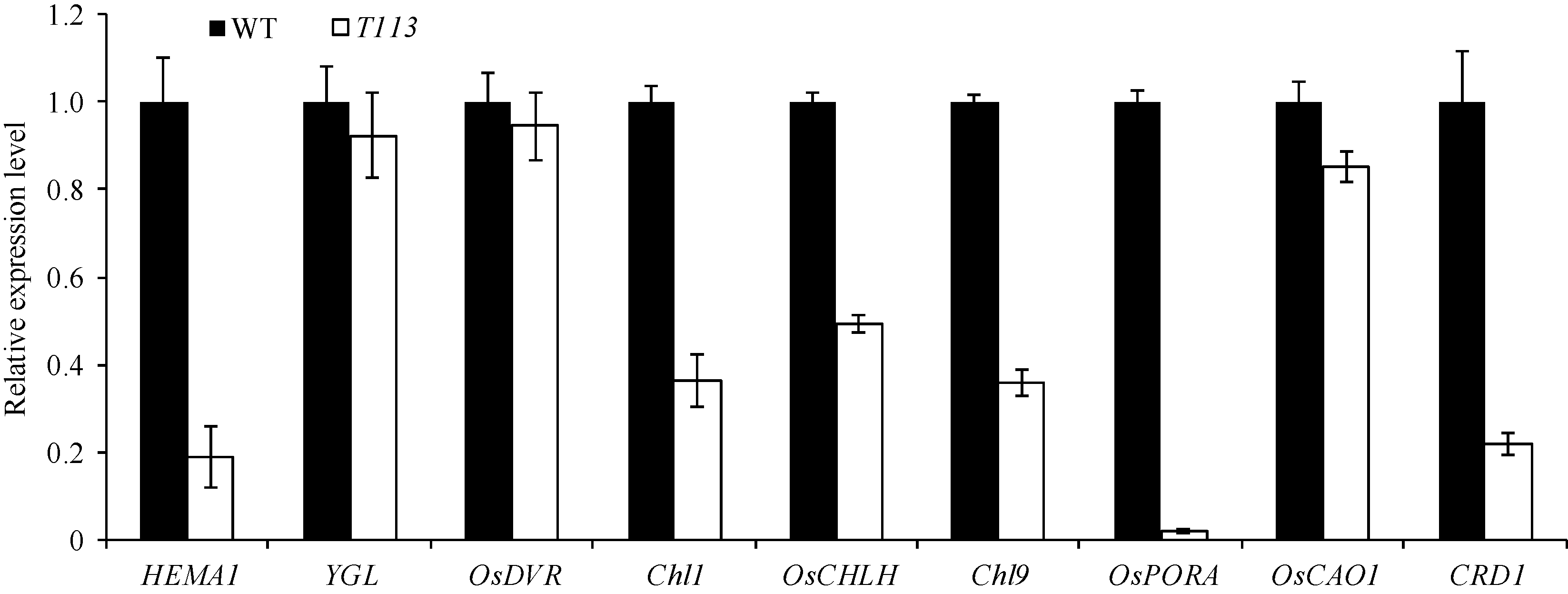
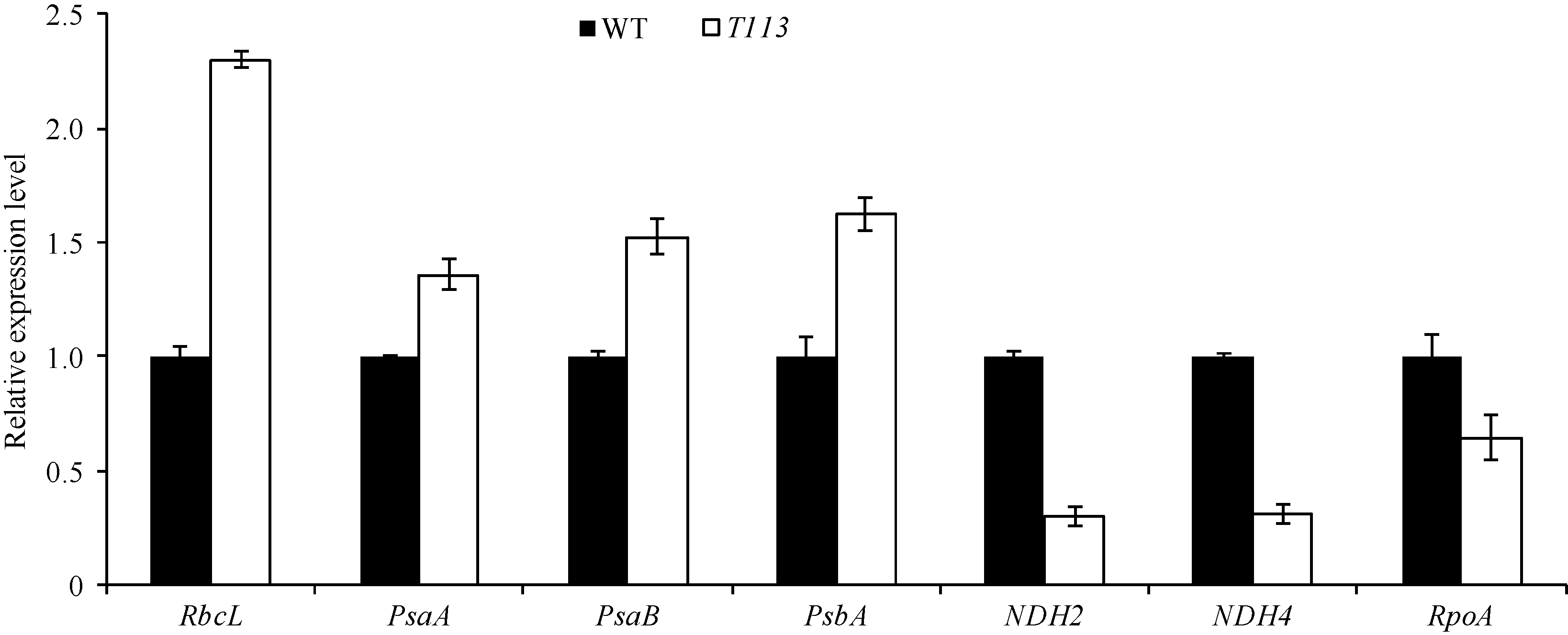
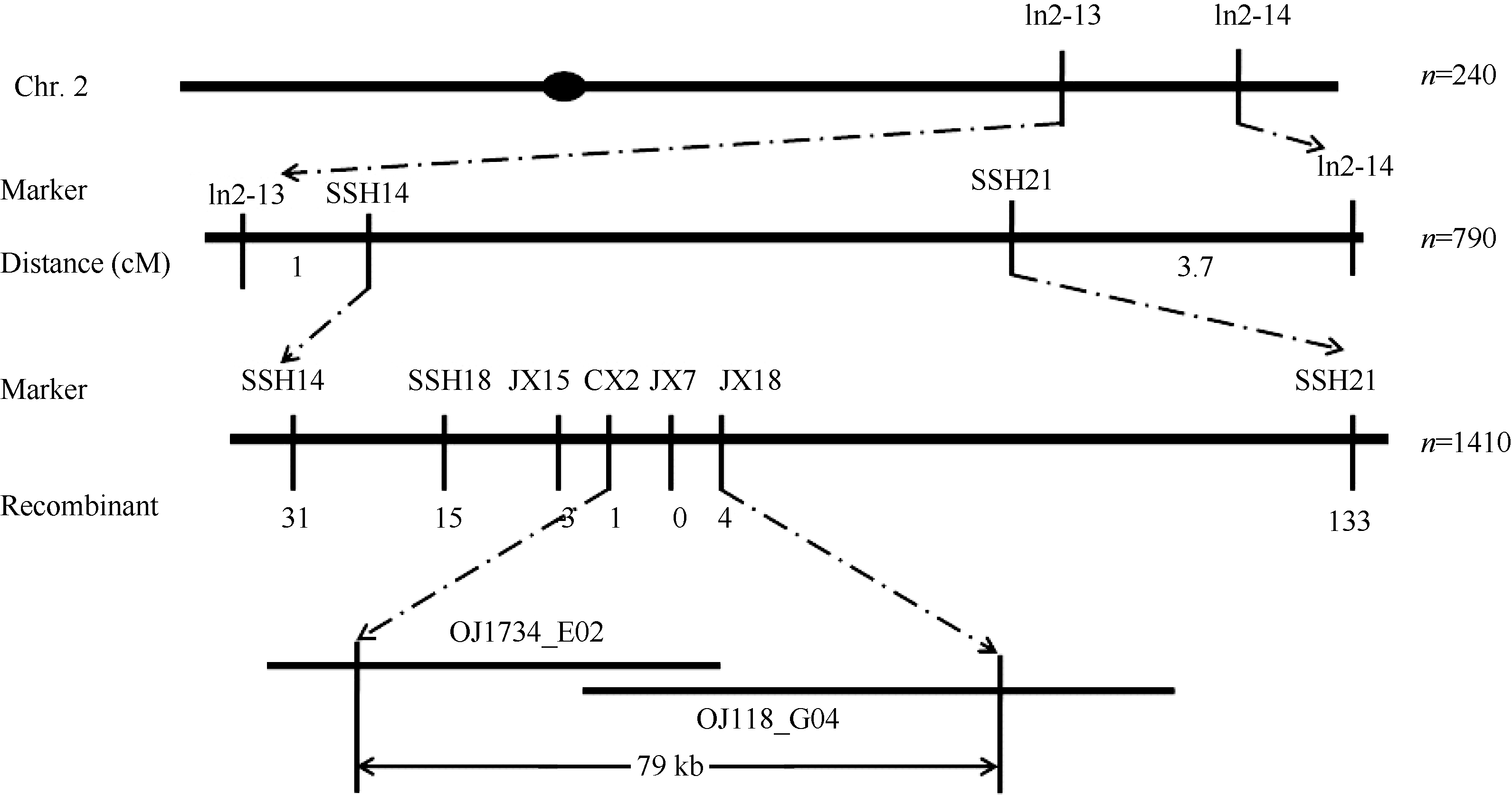
AbstractThe yellow-green leaf mutant T113, which was isolated from a T-DNA mutant pool with Dongjin variety as the background material, showed a yellow-green leaf phenotype in whole developing stage. Compared with wild type, the contents of chlorophyll and carotenoid decreased, the yellow-green leaf became more and more obvious along with developing in T113. At maturity, the number of productive panicles per plant, panicle length, seed setting rate, 1000-grain weight and plant height reduced. The date of heading of T113 also delayed. The phenotype of mutant was not affected by temperature. Ultrastructural analysis showed that the chloroplast of mutant was brighter than that of wild type, the mutant developed loosed thylakoid lamellar structures. The expression of genes associated with chlorophyll biosynthetic and chloroplast development of T113changed a lot. Genetic analysis showed that the yellow-green leaf trait of the T113 mutant was controlled by one pair of recessive nuclear genes. Genetic mapping of the mutant gene was conducted using a F2 mapping population of T113/N22. Finally, the mutant gene was mapped between Indel markers CX2 and JX18 on the long arm of chromosome 2 with physical distance of 79 kb, in which 12 predicted genes had been annotated.
Keyword: Oryza sativaL.; Yellow-green leaf mutant; Genetic analysis; Fine-mapping Show Figures Show Figures
Suzuki JY, Bollivar DW, Bauer CE. Genetic analysis of chlorophyll biosynthesis. Annu Rev Genet, 1997, 31: 61-89[本文引用:1][JCR: 18.115]
[2]
Falbel TG, Meehl JB, Staehelin LA. Severity of mutant phenotype in a series of chlorophyll-deficient wheat mutants depends on light intensity and the severity of the block in chlorophyllsynthesis. Plant Physiol, 1996, 112: 821-832[本文引用:1][JCR: 7.394]
[3]
胡忠, 彭丽萍, 蔡永华. 一个黄绿色的水稻细胞核突变体. 遗传学报, 1981, 8: 256-261HuZ, Peng LP, Cai YH. A yellow-green nucleus mutant of rice. Acta Genet Sin, 1981, 8: 256-261 (in Chinese with English abstract)[本文引用:1]
[4]
ZhaoY, Du LF, Yang SH, Li SC, Zhang YZ. . Chloroplast composition and structure differences in a chlorophyll-reduced mutant of oilseed rape seedlings. Acta Bot Sin, 2001, 43: 877-880[本文引用:1][JCR: 0.599]
[5]
EckhardtU, GrimmB, HortensteinerS. Recent advances in chlorophyll biosynthesis and breakdown in higher plants. Plant Mol Biol, 2004, 56: 1-14[本文引用:1][JCR: 4.072]
Zhang HT, Li JJ, Yoo JH, Yoo SC, Cho SH, Koh HJ, Seo SH, Paek NC. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol, 2006, 62: 325-337. [本文引用:2][JCR: 4.072]
[8]
Jung KH, HurJ, Ryu CH, ChoiY, Chung YY, MiyaoA, HirochikaH, AnG. Characterization of a rice chlorophyll- deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol, 2003, 44: 463-472[本文引用:2][JCR: 4.978]
[9]
Wang PR, Gao JX, Wan CM, Zhang FT, Xu ZJ, Huang XQ, Sun XQ, Deng XJ. Divinyl chlorophyll (ide) a can be converted to monovinyl chlorophyll (ide) a by a divinyl reductase in rice. Plant Physiol, 2010, 153: 994-1003[本文引用:1][JCR: 7.394]
[10]
SakurabaY, Rahman ML, Cho SH, Kim YS, Koi HJ, Yoo SC, Paek NC. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J, 2013, 74: 122-133[本文引用:1][JCR: 6.815]
[11]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40. [本文引用:1][JCR: 7.394]
[12]
LeeS, Kim JH, Yoo ES, Lee CH, HirochikaH, AnG. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol Biol, 2005, 57: 805-818[本文引用:1][JCR: 4.072]
[13]
Terry MJ, Kendrick RE. Feedback inhibition of chlorophyll synthesis in the phytochrome chromophore-deficient aurea and yellow-green-2 mutants of tomato. Plant Physiol, 1999, 119: 143-152[本文引用:2][JCR: 7.394]
[14]
Lopez-JuezE. Plastid biogenesis between light and shadows. J Exp Bot, 2007, 58: 11-26[本文引用:1][JCR: 5.794]
[15]
SakamotoW, Miyagishima SY, JarvisP. Chloroplast biogenesis: control of plastid development, protein import, division and inheritance. Arabidopsis Book, 2008, 6: e110[本文引用:1]
[16]
Webber AN, MalkinR. Photosystem I reaction-centre proteins contain leucine zipper motifs. A proposed role in dimer formation. FEBS Lett, 1990, 264: 1-4[本文引用:1][JCR: 3.341]
AnderssonI, BacklundA. Structure and function of Rubisco. Plant Physiol Biochem, 2008, 46: 275-291[本文引用:1]
[19]
Peng LW, YamamotoH, ShikanaiT. Structure and biogenesis of the chloroplast NAD(P)H dehydrogenase complex. Biochim Biophys Acta, 2011, 1807: 945-953[本文引用:1]
[20]
Lichtenthaler HK. Chlorophylls and carotenoids: pigments of photosynthetic membranes. Methods Enzymol, 1987, 148: 350-382[本文引用:1][JCR: 2.194]
[21]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C (T)) method. Methods, 2001, 25: 402-408[本文引用:1][JCR: 3.221]
Shi YF, ChenJ, Liu WQ, Huang QN, ShenB, LeungH, Wu JL. Genetic analysis and gene mapping of a new rolled-leaf mutant in rice (Oryza sativa L. ). Sci China C Life Sci, 2009, 52: 885-890[本文引用:1][CJCR: 0.478]
[24]
LanT, WangB, Ling QP, Xu CH, Tong ZJ, Liang KJ, Duan YL, JinJ, Wu WR. Fine mapping of cisc(t), a gene for cold-induced seedling chlorosis, and identification of its cand idate in rice. Chin Sci Bull, 2010, 55: 3149-3153[本文引用:1][CJCR: 1.541]
[25]
Agrawal GK, YamazakiM, KobayashiM, HirochikaR, MiyaoA, HirochikaH. Screening of the rice viviparous mutants generated by endogenous retrotransposon Tos17 insertion tagging of a zeaxanthin epoxidase gene and a novel OsTATC gene. Plant Physiol, 2001, 125: 1248-1257[本文引用:1][JCR: 7.394]
[26]
Von Gromoff ED, AlawadyA, MeineckeL, GrimmB, Beck CF. Heme, a plastid-derived regulator of nuclear gene expression in Chlamydomonas. Plant Cell, 2008, 20: 552-567[本文引用:1][JCR: 9.575]
[27]
LeeS, RyooN, Jeon JS, Guerinot ML, AnG. Activation of rice Yellow Stripe1-Like 16 (OsYSL16) enhances iron efficiency. Mol Cells, 2012, 33: 117-126[本文引用:1][JCR: 2.242]
[28]
KakeiY, IshimaruY, KobayashiT, YamakawaT, NakanishiH, Nishizawa NK. OsYSL16 plays a role in the allocation of iron. Plant Mol Biol, 2012, 79: 583-594[本文引用:1][JCR: 4.072]
[29]
HudsonD, Guevara DR, Hand AJ, Xu ZH, Hao LX, ChenX, ZhuT, Bi YM, Rothstein SJ. Rice Cytokinin GATA Transcription Factor1 regulates chloroplast development and plant architecture. Plant Physiol, 2013, 162: 132-144[本文引用:1][JCR: 7.394]
[30]
Liu WZ, Fu YP, Hu GC, Si HM, ZhuL, WuC, Sun ZX. Identification and fine mapping of a thermo-sensitive chlorophyll deficient mutant in rice (Oryza sativa L. ). Planta, 2007, 226: 785-795[本文引用:1][JCR: 3.376]
[31]
李燕群, 钟萍, 高志艳, 朱柏羊, 陈丹, 孙昌辉, 王平荣, 邓晓建. 水稻斑马叶突变体zebra524的表型鉴定及候选基因分析. 中国农业科学, 2014, 47: 2907-2915Li YQ, ZhongP, Gao ZY, Zhu BY, ChenD, Sun CH, Wang PR, Deng XJ. Morphological characterization and cand idate gene analysis of zebra leaf mutant zebra524 in rice. Sci Agric Sin, 2014, 47: 2907-2915 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4]
[32]
Gao QS, Yang ZF, ZhouY, Yin ZT, QiuJ, Liang GH, Xu CW. Characterization of an Abc1 kinase family gene OsABC1-2 conferring enhanced tolerance to dark-induced stress in rice. Gene, 2012, 498: 155-163[本文引用:1][JCR: 2.082]
[33]
ChappellJ. The biochemistry and molecular biology of isoprenoid metabolism. Plant Physiol, 1995, 107: 1-6[本文引用:1][JCR: 7.394]
[34]
Riley MV, Peters MI. The localization of the anion-sensitive ATPase activity in corneal endothelium. Biochim Biophys Acta, 1981, 644: 251-256[本文引用:1]
[35]
SharmaR, PatelV, KrishnaH. Relationship between light, fruit and leaf mineral content with albinism incidence in strawberry (Fragaria × ananassa Duch. ). Sci Hortic, 2006, 109: 66-70[本文引用:1][JCR: 0.92]
[36]
BroughtonS. Ovary co-culture improves embryo and green plant production in anther culture of Australian spring wheat (Triticum aestivum L. ). Plant Cell Tiss Org Cult, 2008, 95: 185-195[本文引用:1]
[37]
Xu ZJ, NakajimaM, SuzukiY, YamaguchiI. Cloning and characterization of the abscisic acid-specific glucosyltransferase gene from adzuki bean seedlings. Plant Physiol, 2002, 129: 1285-1295[本文引用:1][JCR: 7.394]
[38]
Bae JH, Sohn JH, Park CS, Rhee JS, Choi ES. Cloning and functional characterization of the SUR2/SYR2 gene encoding sphinganine hydroxylase in Pichia ciferrii. Yeast, 2004, 21: 437-443[本文引用:1]
 , 万建民
, 万建民







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}