关键词:陆地棉; 枯萎病; Solexa; 基因表达谱 Expression Profiling Analysis between Resistant and Susceptible Cotton Cultivars ( Gossypium hirsutum L.) in Response to Fusarium Wilt HAN Ze-Gang, ZHAO Zeng-Qiang, LI Hui-Hui, ZHANG Xi, LI Xiao-Ling, ZHANG Wei* Agricultural College of Shihezi University / Laboratory of Cotton Molecular Breeding, Shihezi 832000, China
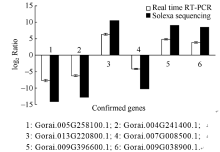
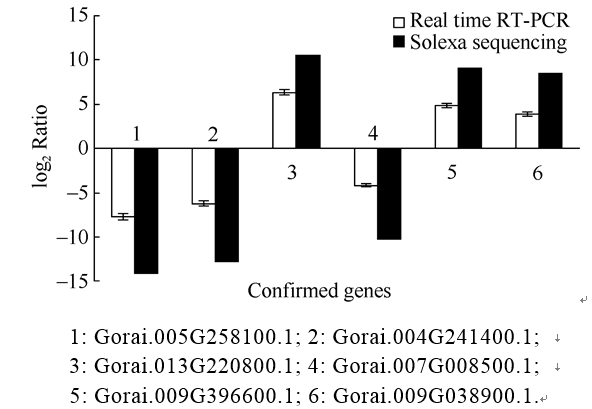
AbstractFusarium Wilt resistant cotton cultivar Zhongmiansuo 12 and susceptible cultivar Xinluzao 7 were used to analyze the gene expression profiling of root tissues at three hours and six hours after the seedlings infection by Fusarium Wilt using Solexa sequencing technology. Compared with no infection, 4447 and 5481 differential genes after three hours and six hours infected by Fusarium Wilt were identified, and compared six hours with three hours, 2559 differential genes were identified in Zhongmiansuo 12; while 8615, 6727, and 2078 was respectively identified in Xinluzao 7. There were 1879 and 500 differential genes in three hours and six hours after infection when the two cultivars were compared. According to the Gene Ontology, these genes were divided into these groups of biological process, cellular component and molecular function; then further subdivided into 48 functional categories. By analyzing the pathways, the most of them were identified in 6 h/0 h after infection between Zhongmiansuo 12 and Xinluzao 7, which were 126 each in the two cultivars; the least of pathways were at 6 h after infection in two cultivars, which was 89 only. However, all the pathways in each comparison group could be classified into 13 categories, such as biosynthesis of other secondary metabolites, glycan biosynthesis and metabolism, environmental adaptation, and immune system. A pathway, named plant-pathogen interaction in the environmental adaptation and immune system category, was involved in 996 differential genes; and the number of up regulated genes was 444 and that of down regulated genes was 552. The most differential genes were in WRKY transcription factor family, the serine/threonine kinase had the medium number of differential genes, while DNA damage-repair/toleration protein, JAZ1, RAR1, RPM1-interacting protein, S locus specific glycoprotein S6 precursor and caltractin had the least genes. Finally, six random genes were subjected to RT-PCR and the results validated the Solexa sequencing conclusions.
Keyword: Gossypium hirsutum; Fusarium wilt; Solexa; Expression profiling Show Figures Show Figures
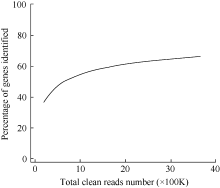
图1 Z-3的Solexa测序饱和度分析Fig. 1 Solexa sequencing saturation analysis of Z-3
2.2 差异表达基因统计分析经Solexa测序后, 对差异表达基因统计分析(表2)表明, 2个品种在枯萎病菌侵染后的3 h和6 h, 分别与对照相比, 差异表达基因中的下调基因均明显多于上调基因, 且感病品种的差异表达基因明显多于抗病品种。在中棉所12中, 与对照(Z-0)相比, 枯萎病菌侵染后3 h (Z-3)差异表达基因数目为4447个。其中上调基因数目为1300个, 下调基因数目为3147个。枯萎病菌处理后6 h (Z-6), 与对照相比, 差异表达基因数目为5481个。其中上调基因数目为1494个, 下调基因数目为3987个。而Z-6与Z-3相比, 差异表达基因的数目较少, 为2559个。其中上调基因数目为1020个, 下调基因数目为1539个。在新陆早7号中, 与对照(X-0)相比, 枯萎病菌侵染后3 h (X-3)的差异表达基因数目为8615个。上调基因数目为2118个, 下调基因数目为6497个。枯萎病菌处理后6 h (X-6)与X-0相比, 差异表达基因数目为6727个。上调基因数目为2038个, 下调基因数目为4689个。而X-6与X-3相比, 差异表达基因的数目为2078个。其中上调基因数目为1010个, 下调基因数目为1068个。 比较2个品种在枯萎病菌侵染同一时间点的差异表达基因(表2)表明, 下调基因数目依然多于上调基因, 但是差异表达基因的总数迅速下降。其中, 在受到枯萎病菌侵染后的3 h (Z-3 vs. X-3), 抗病品种相对于感病品种的差异表达基因数目为1879个。上调基因数目为874个, 下调基因数目为1005个。而在受到枯萎病菌侵染后的6 h (Z-6 vs. X-6), 抗病品种相对于感病品种的差异表达基因数目为500个。上调基因数目为314个, 下调基因数目为186个。 表2 Table 2 表2(Table 2)
表2 差异表达基因 Table 2 Number of differentially expressed genes
比对组 Contrasting group
差异表达基因 Number of differentially expressed genes
上调基因 Up-regulated
下调基因 Down-regulated
已知(百分比)a Known (percentage)a
未知(百分比)a Unknown (percentage)a
已知(百分比)a Known (percentage)a
未知(百分比)a Unknown (percentage)a
Z-3 vs. Z-0
4447
865(19.45%)
435(9.78%)
2209(49.67%)
938(21.09%)
Z-6 vs. Z-0
5481
1076(19.63%)
418(7.63%)
2947(53.77%)
1040(18.97%)
Z-6 vs. Z-3
2559
717(28.02%)
303(11.84%)
885(34.58%)
654(25.56%)
X-3 vs. X-0
8615
1398(16.23%)
720(8.36%)
4732(54.93%)
1765(20.49%)
X-6 vs. X-0
6727
1351(20.08%)
687(10.21%)
3633(54.01%)
1056(15.70%)
X-6 vs. X-3
2078
661(31.81%)
349(16.79%)
771(37.10%)
297(14.29%)
Z-3 vs. X-3
1879
596(31.72%)
278(14.80%)
755(40.18%)
250(13.30%)
Z-6 vs. X-6
500
188(37.60%)
126(25.20%)
153(30.60%)
33(6.60%)
a Known stands for genes with Gene Ontology annotation; Unknown stands for genes without annotation. a已知基因为Gene Ontology中有注释的基因; 未知基因为Gene Ontology中没有注释的基因。
表2 差异表达基因 Table 2 Number of differentially expressed genes
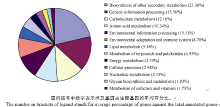
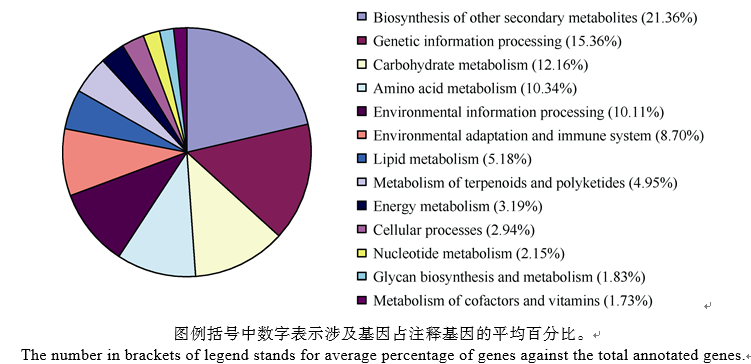
2.3 差异表达基因GO功能分析GO分析将104 610个差异表达基因分为3个功能注释本体, 分别描述基因的生物学过程、细胞组分和分子功能。中棉所12比对组Z-3 vs. Z-0、Z-6 vs. Z-0和Z-6 vs. Z-3分别有15 578、21 581和9683个, 而新陆早7号比对组分别有33 462、27 728和7377个unigene至少含有一种功能注释。2个品种相同处理时间的比对组Z-3 vs. X-3和Z-6 vs. X-6分别有7052个和1750个unigene至少含有一种功能注释。生物学过程、细胞组分和分子功能又可被进一步划分为48个类别(表3), 依次为23、14和11个类别。其中催化活性, 代谢过程和蛋白结合注释到的差异表达基因最多, 分别为12 397、12 158和11 678个, 而翻译调控活性类别最少, 仅有1个差异表达基因。 表3 Table 3 表3(Table 3)
表3 差异表达基因的功能分类 Table 3 Functional categorization of the differentially expressed genes
功能类别 Class
基因数量The number of genes
合计 Total
Z-3 vs. Z-0
Z-6 vs. Z-0
Z-6 vs. Z-3
X-3 vs. X-0
X-6 vs. X-0
X-6 vs. X-3
Z-3 vs. X-3
Z-6 vs. X-6
生物学过程Biological process
代谢过程 Metabolic process
1560
2139
977
3155
2686
773
703
165
12158
细胞过程 Cellular process
1333
1822
869
2686
2273
643
578
144
10348
刺激反应 Response to stimulus
682
913
429
1244
1085
333
316
91
5093
单生物过程 Single-organism process
633
870
418
1345
1083
272
282
71
4974
定位 Localization
341
470
218
648
575
149
161
43
2605
定位建立 Establishment of localization
332
456
213
630
557
147
158
43
2536
生物调节 Biological regulation
286
407
194
610
507
126
126
33
2289
发育过程 Developmental process
232
337
170
585
471
122
127
26
2070
生物过程调控 Regulation of biological process
208
318
147
487
402
95
103
28
1788
细胞组分组织生物合成 Cellular component organization or biogenesis
吴征彬, 杨业华, 刘小丰, 王强. 枯萎病对棉花产量和纤维品质的影响. 棉花学报, 2004, 16: 236-239Wu ZB, Yang YH, Liu XF, WangQ. Effect of Fusarium wilt on the cotton yield and fiber quality. Cott Sci, 2004, 16: 236-239 (in Chinese with English abstract)[本文引用:1]
[2]
徐秋华, 张献龙, 聂以春, 冯纯大. 我国棉花抗枯萎病品种的遗传多样性分析. 中国农业科学, 2002, 35: 272-276Xu QH, Zhang XL, Nie YC, Feng CD. Genetic diversity evaluation of cultivars (G. hirsumtum L. ) resistant to Fusarium wilt by RAPD markers. Sci Agric Sin, 2002, 35: 272-276 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4]
[3]
刘坤. 海岛棉ERF族B3和B1亚组转录因子基因的克隆与特征分析. 中国农业科学院硕士学位论文, 北京, 2011. pp 26-30LiuK. Gene Cloning and Characterization of B3 and B1 Subgroup Transcription Factors of ERF Family from Gossypium barbadense L. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2011. pp 26-30 (in Chinese with English abstract)[本文引用:1]
[4]
韩庆典, 陈志伟, 段远霖, 兰涛, 官华忠, 周元昌, 吴为人. 利用基因芯片检测水稻细菌性条斑病抗性相关基因. 分子植物育种, 2008, 6: 239-244Han QD, Chen ZW, Duan YL, LanT, Guan HZ, Zhou YC, Wu WR. Detection of genes related to resistance to bacterial leaf streak in rice using microarray. Mol Plant Breed, 2008, 6: 239-244 (in Chinese with English abstract)[本文引用:1][CJCR: 0.805]
[5]
何兰兰, 柴蒙亮, 韩泽刚, 赵曾强, 张薇. 棉花抗枯萎病相关ERF-B3亚组转录因子的克隆与表达. 西北植物学报, 2013, 33: 2375-2381He LL, Chai ML, Han ZG, Zhao ZQ, ZhangW. Cloning and expression of ERF-B3 subgroup transcription factor related to resistance Fusarium Wilt in cotton. Acta Bot Boreali-Occident Sin, 2013, 33: 2375-2381 (in Chinese with English abstract)[本文引用:1][CJCR: 0.947]
[6]
谢为博. 基于表达谱芯片和新一代测序技术的高通量基因分型方法的开发. 华中农业大学博士学位论文, 湖北 武汉, 2010. pp 17-18Xie WB. Development of High Throughput Genotyping Methods Based on DNA Microarray and New Generation Sequencing Technologies. PhD Dissertation of Huazhong Agricultural University, Wuhan, China, 2010. pp 17-18 (in Chinese with English abstract)[本文引用:1]
[7]
WuJ, Zhang YL, Zhang HQ, HuangH, Folta KM, LuJ. Whole genome wide expression profiles of Vitis amurensis grape responding to downy mildew by using Solexa sequencing technology. BMC Plant Biol, 2010, 10: 234[本文引用:1][JCR: 3.942]
[8]
Bai TT, Xie WB, Zhou PP, Wu ZL, Xiao WC, ZhouL, SunJ, Ruan XL, Li HP. Transcriptome and expression profile analysis of highly resistant and susceptible banana roots challenged with Fusarium oxysporum f. sp. cubense tropical race 4. PloS One, 2013, 8: e73945[本文引用:1][JCR: 3.534]
[9]
Yu SC, Zhang FL, Yu YJ, Zhang DS, Zhao XY, Wang WH. Transcriptome profiling of dehydration stress in the Chinese cabbage (Brassica rapa L. ssp. pekinensis) by tag sequencing. Plant Mol Bio Rep, 2012, 30: 17-28[本文引用:1]
[10]
PangT, Ye CY, Xia XL, Yin WL. De novo sequencing and transcriptome analysis of the desert shrub, Ammopiptanthus mongolicus, during cold acclimation using Illumina/Solexa. BMC Gene, 2013, 14: 488-503[本文引用:1]
[11]
Shan XH, Li YD, JiangY, Jiang ZL, Hao WY, Yuan YP. Transcriptome profile analysis of maize seedlings in response to high-salinity, drought and cold stresses by deep sequencing. Plant Mol Biol Rep, 2013, 31: 1485-1491[本文引用:1][JCR: 2.374]
[12]
Andrew JC, Liu DC, RamilM, Yue I C H, Rachid S. Transcriptome profiling of leaf elongation zone under drought in contrasting rice cultivars. PloS One, 2013, 8: e54537[本文引用:1][JCR: 3.534]
[13]
Chen JH, Song YP, ZhangH, Zhang DQ. Genome-wide analysis of gene expression in response to drought stress in Populus simonii. Plant Mol Biol Rep, 2013, 31: 946-962[本文引用:1][JCR: 2.374]
[14]
WangY, XuL, Chen YL, ShenH, Gong YQ, CeciliaL, Liu LW. Transcriptome profiling of radish (Raphanus sativus L. ) root and identification of genes involved in response to lead (Pb) stress with next generation sequencing. PloS One, 2013, 8: e66539[本文引用:1][JCR: 3.534]
[15]
彭姗, 吕学莲, 高峰, 李国英, 李晖. 一种新的棉花黄, 枯萎病快速接种方法的研究. 棉花学报, 2008, 20: 174-178PengS, Lü XL, GaoF, Li GY, LiH. Study on a new rapid inoculation method for verticillium wilt and Fusarium wilt of cotton. Cott Sci, 2008, 20: 174-178 (in Chinese with English abstract)[本文引用:1]
[16]
胡根海, 喻树迅. 利用改良的CTAB法提取棉花叶片总RNA. 棉花学报, 2007, 19: 69-70Hu GH, Yu SX. Extraction of high-quality total RNA in cotton leaf with improved CTAB method. Cotton Sci, 2007, 19: 69-70 (in Chinese with English abstract)[本文引用:1][CJCR: 1.318]
[17]
MortazaviA, Williams BA, McCue K, Schaeffer L, Wold B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Natl Methods, 2008, 5: 621-628[本文引用:1]
[18]
AudicS, Claverie JM. The significance of digital gene expression profiles. Genome Res, 1997, 7: 986-995[本文引用:1][JCR: 13.852]
王刚. 棉花幼苗盐胁迫条件下Solexa转录组测序结果的分析及验证. 山东农业大学硕士学位论文, 山东 泰安, 2011. pp 36-38WangG. Transcription Analysis of Young Cotton (Gossypium hirsutum) Seeding under Salt Stress via Solexa Sequencing. MS Thesis of Shand ong Agricultural University, Tai’an, China, 2011. pp 36-38 (in Chinese with English abstract)[本文引用:1]
[21]
陈欢, 杨美英, 王真慧, 孙旸, 王刚, 魏玮, 关瑜, 王法微, 李海燕, 陈光. 大豆籽粒不同发育时期基因表达谱分析. 中国油料作物学报, 2012, 34: 129-135ChenH, Yang MY, Wang ZH, SunY, WangG, WeiW, GuanY, Wang FW, Li HY, ChenG. Gene expression profile of developing soybean seeds. Chin J Oil Crop Sci, 2012, 34: 129-135 (in Chinese with English abstract)[本文引用:2][CJCR: 1.087]
[22]
Wei MM, Song MZ, Fan SL, Yu SX. Transcriptomic analysis of differentially expressed genes during anther development in genetic male sterile and wild type cotton by digital gene-expression profiling. BMC Genetics, 2013, 14: 97[本文引用:1][JCR: 2.356]
[23]
罗红丽, 陈银华. 植物抗病反应相关转录因子的研究进展. 热带生物学报, 2011, 2: 83-88Luo HL, Chen YH. Advance on transcription factors involved in plant disease resistance response. Chin J Trop Crops, 2011, 2: 83-88 (in Chinese with English abstract)[本文引用:2][CJCR: 0.618]
[24]
郭艳玲, 张鹏英, 郭默然, 陈靠山. 次生代谢产物与植物抗病防御反应. 植物生理学报, 2012, 48: 429-434Guo YL, Zhang PY, Guo MR, Chen KS. Secondary metabolites and plant defence against pathogenic disease. Plant Physiol J, 2012, 48: 429-434 (in Chinese with English abstract)[本文引用:1][JCR: 2.77]
[25]
Pieterse C MJ, Leon-ReyesA, Van der EntS, Van Wees S C M. Networking by small-molecule hormones in plant immunity. Nat Chem Biol, 2009, 5: 308-316[本文引用:1][JCR: 13.217]
[26]
Anderson JP, BadruzsaufariE, Schenk PM, Manners JM, Desmond OJ, EhlertC, Maclean DJ, Ebert PR, KazanK. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell, 2004, 16: 3460-3479[本文引用:1][JCR: 9.575]
[27]
张春宝, 赵丽梅, 赵洪锟, 董英山. 植物蛋白激酶研究进展. 生物技术通报, 2011, (10): 17-23Zhang CB, Zhao LM, Zhao HK, Dong YS. Advances in plant protein kinase. Biol Bull, 2011, (10): 17-23 (in Chinese with English abstract)[本文引用:1][JCR: 1.567]
[28]
Song WY, Wang GL, Chen LL. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science, 1995, 270: 1804-1806[本文引用:1]
[29]
周庆红, 李成琼, 匡全. 植物蛋白激酶研究进展. 生物学杂志, 2003, 20(3): 1-4Zhou QH, Li CQ, KuangQ. Advances in plant protein kinase. J Biol, 2003, 20(3): 1-4 (in Chinese with English abstract)[本文引用:1][CJCR: 0.56]
[30]
't Hoen P A C, AriyurekY, Thygesen HH, VreugdenhilE, Vossen RH, de Menezes RX, Boer JM, van Ommen GB, den Dunnen JT. Deep sequencing-based expression analysis shows major advances in robustness, resolution and inter-lab portability over five microarray platforms. Nucl Acids Res, 2008, 36: e141[本文引用:1]


{kind=link}
{kind=link}
{kind=link}