关键词:甘蓝型油菜; 全基因组关联分析; SNP; 株高; 候选基因 Genome-wide Association Analysis of Plant Height in Rapeseed ( Brassica napus) TANG Min-Qiang1, CHENG Xiao-Hui1, TONG Chao-Bo1,2, LIU Yue-Ying1, ZHAO Chuan-Ji1, DONG Cai-Hua1,2, YU Jing-Yin1, MA Xiao-Gen1, HUANG Jun-Yan1,2,*, LIU Sheng-Yi1,2 1Oil Crops Research Institute, Chinese Academy of Agricultural Sciences / Key Laboratory of Biology and Genetic Improvement of Oil Crops, Ministry of Agriculture, Wuhan 430062, China
2Hubei university / Hubei Collaborative Innovation Center for Green Transformation of Bio-Resources, Wuhan 430062, China
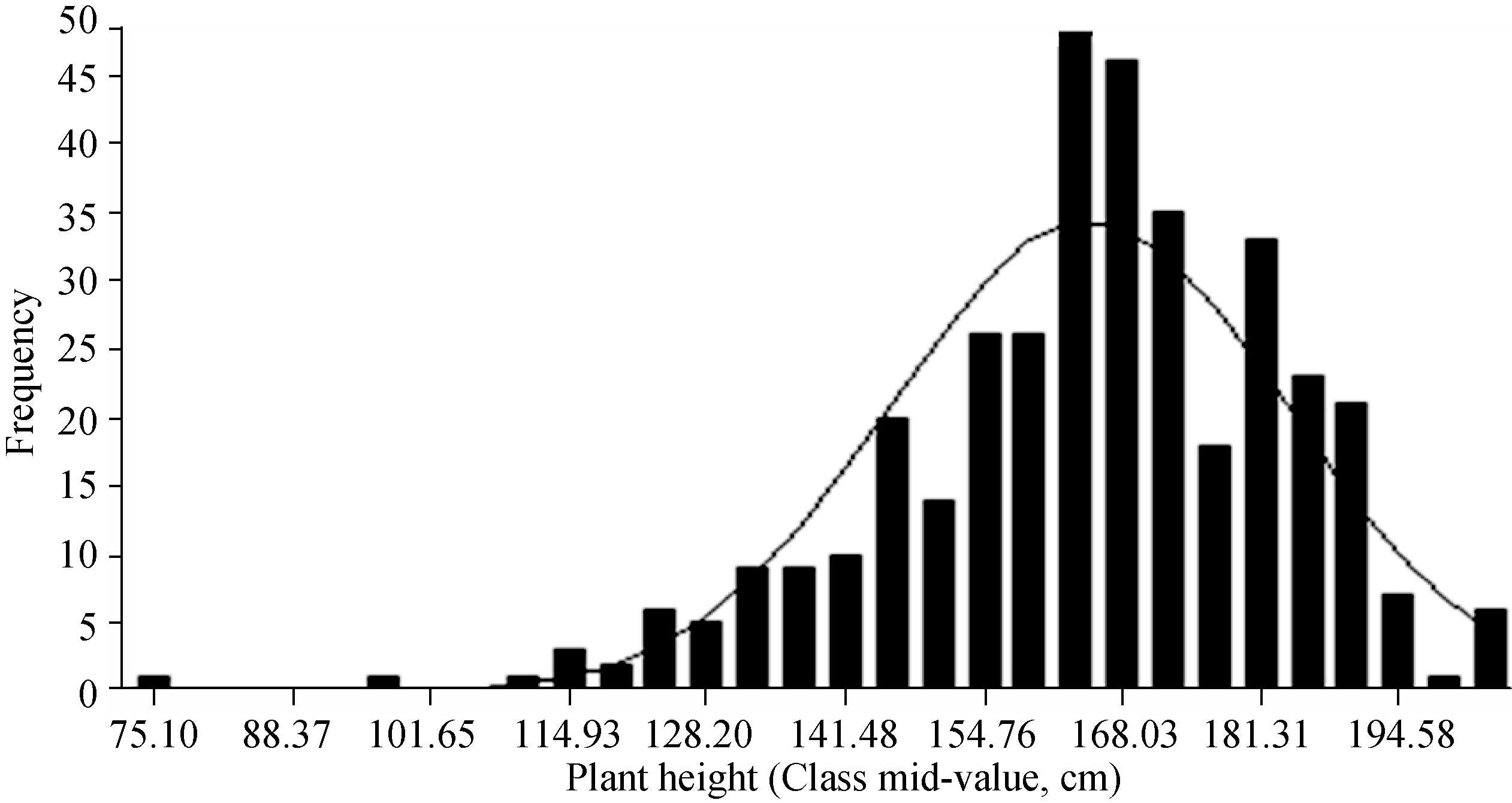
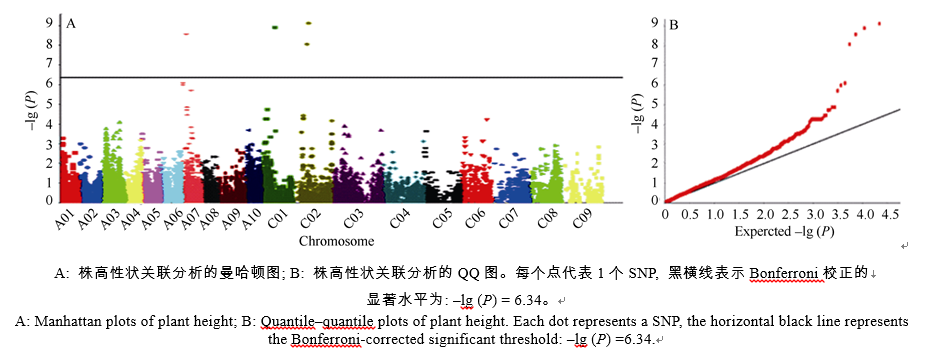
AbstractAppropriate plant height is a trait related to lodging resistance and mechanical harvest, and ideal plant height is a basis of ideotype for high yield. The plant height of 371 rapeseed accessions collected from different growing regions in the world was measured in field experiments, all accessions were genotyped using a 60K SNP chip and 21856 informative SNPs were acquired, then the data of SNPs and plant height were used for genome-wide association analysis. Major results were as follows: 1) 4 significant SNPs related to plant height were detected, including Bn-A07-p3583161, Bn-scaff_22790_1-p1271170, Bn-scaff_ 20735_1-p42779, and Bn-scaff_18702_1-p589589, which located in A07, C01 and C02 chromosomes, and explained 11.33%, 11.75%, 12.31%, and 10.97% of phenotypic variation, respectively; 2) According to the reference genome annotation information, three candidate genes corresponding to the first three significant SNPs were predicted, each gene was located in 492.0, 9.5, and 69.5 kb away from the three SNPs, respectively ; 3) The significant (two-tailed t-test) difference of plant height between means of two allelic groups of accessions corresponding to each significant SNP was 11.09, 15.12, 10.48, and 8.93 cm, respectively.
Keyword: Brassica napus L.; Genome-wide association analysis; SNP; Plant height; Candidate gene Show Figures Show Figures
王宇. 甘蓝型油菜矮秆突变体M176的株高性状遗传分离分析和QTL分析. 南京农业大学硕士学位论文, 江苏南京, 2012. pp 6-10WangY. Genetic Segregation Analysis and QTL Mapping of The Plant Height Trait in Brassica napus L. MS Thesis of Nanjing Agricultural University, Nanjing, China, 2012. pp 6-10 (in Chinese with English abstract)[本文引用:2]
[2]
李加纳. 我国油菜生产加工现状、面临的挑战及对策. 北京: 中国农业出版社, 2002. pp 27-31Li JN. The Status Quo of Chinese Rapeseed Production, Processing, Challenges and Strategies. Beijing: China Agriculture Press, 2002. pp 27-31(in Chinese)[本文引用:1]
[3]
Becker HC, Loptien H. RobblenG. Breeding of Brassica: an overview. In: Gomez-Campo C, ed. Biology of Brassica coenospecies. Amsterdam: Elsevier, Science BV, 1999. pp 413-460[本文引用:1]
FoissetN, DeloumreR, BarretP, RenardM. Molecular tagging of the dwarf BREIZH (Bzh) gene in Brassica napus. Theor Appl Genet, 2006, 91: 756-761[本文引用:1][JCR: 3.658]
[6]
浦惠明, 戚存扣, 傅寿仲. 甘蓝型矮秆油菜矮源1号鉴定研究初报. 中国种业, 1995, (1): 23-24Pu HM, Qi CK, Fu SZ. The identified preliminary report of the dwarf source 1 in Brassica napus L. China Seed Ind, 1995, (1): 23-24 (in Chinese)[本文引用:1][CJCR: 0.3657]
[7]
黄天带. 甘蓝型油菜矮秆突变体及矮秆基因分子标记的研究. 华中农业大学硕士学位论文, 湖北武汉, 2003. pp 1-5Huang TD. Studies on a Dwarf Mutant and Molecular Markers of Dwarf Gene in Brassica napus L. MS Thesis of Huazhong Agricultural University, Wuhan, China, 2003. pp 1-5 (in Chinese with English abstract)[本文引用:1]
[8]
张倩. 甘蓝型油菜主要株型性状的遗传分析和QTL初步定位. 西南大学硕士学位论文, 重庆, 2013. pp 3-7ZhangQ. Genetic Effects Analysis and QTL Mapping of Major Plant-Type Traits in Brassica napus L. MS Thesis of Southwest University, Chongqing, China, 2013. pp 3-7 (in Chinese with English abstract)[本文引用:1]
[9]
ShiJ, LiR, ZouJ, LongY, MengJ. A dynamic and complex network regulates the heterosis of yield-correlated traits in rapeseed (Brassica napus L. ). PLoS One, 2011, 6: e21645[本文引用:2][JCR: 3.73]
[10]
Basunand aP, RadoevM, EckeW, FriedtW, Becker HC, Snowdon RJ. Comparative mapping of quantitative trait loci involved in heterosis for seedling and yield traits in oilseed rape (Brassica napus L. ). Theor Appl Genet, 2010, 120: 271-281[本文引用:2][JCR: 3.658]
[11]
ChenW, ZhangY, LiuX, ChenB, TuJ, Fu TD. Detection of QTL for six yield-related traits in oilseed rape (Brassica napus) using DH and immortalized F2 populations. Theor Appl Genet, 2007, 115: 849-858[本文引用:2][JCR: 3.658]
[12]
Butruille DV, Guries RP, Osborn TC. Linkage analysis of molecular markers and quantitative trait loci in populations of inbred backcross lines of Brassica napus L. Genetics. 1999, 153: 949-964[本文引用:2][JCR: 4.389]
[13]
Quijada PA, Udall JA, LambertB, Osborn TC. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L. ): 1. Identification of genomic regions from winter germplasm. Theor Appl Genet, 2006, 113: 549-561[本文引用:2][JCR: 3.658]
[14]
Udall JA, Quijada PA, LambertB, Osbom TC. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L. ): 2. Identification of alleles from unadapted germplasm. Theor Appl Genet, 2006, 113: 597-609[本文引用:3][JCR: 3.658]
[15]
Zhao JY, Becker HC, Ding HD, Zhang YF, Zhang DQ, EckeW. QTL of three agronomically important traits and their interactions with environment in a European × Chinese rapeseed population. Acta Genet Sin, 2005, 32: 969-978[本文引用:2]
[16]
Shi JQ, Li RY. Unraveling the complex trait of crop yield with quantitative trait loci mapping in Brassica napus. Genetics, 2009, 182: 851-861[本文引用:2][JCR: 4.389]
[17]
杨小红, 严建兵, 郑艳萍. 植物数量性状关联分析研究进展. 作物学报, 2007, 33: 523-530Yang XH, Yan JB, Zheng YP. Reviews of association analysis for quantitative traits in plants. Acta Agron Sin, 2007, 33: 523-530 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[18]
TianF, Bradbury PJ, Brown PJ, HungH, SunQ, Flint-GarciaS, Rocheford TR, McMullen M D, Holland J B, Buckler E S. Genome-wide association study of leaf architecture in the maize nested association mapping population. Nat Genet, 2011, 159-162[本文引用:1][JCR: 35.209]
[19]
ZhaoK, Tung CW, Eizenga GC, Wright MH, Ali ML, Price AH, Norton GJ, Islam MR, ReynoldsA, MezeyJ, McClung A M, Bustamante C D, McCouch S R. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat Commun, 2011, 2: 467[本文引用:1][JCR: 10.015]
[20]
AtwellS, Huang YS, Vilhjálmsson BJ, WillemsG, HortonM, LiY, MengD, PlattA, Tarone AM, Hu TT, JiangR, Muliyati NW, ZhangX, Amer MA, BaxterI, BrachiB, ChoryJ, DeanC, DebieuM, MeauxJ, Ecker JR, FaureN, Kniskern JM, Jones JD, MichaelT, NemriA, RouxF, Salt DE, TangC, TodescoM, Traw MB, WeigelD, MarjoramP, Borevitz JO, BergelsonJ, NordborgM. Genome-wide association study of 10phenotypes in a common set of Arabidopsis thaliana inbred lines. Nature, 2010, 465: 627-631[本文引用:1][JCR: 38.597]
[21]
Harper AL, TrickM, HigginsJ, FraserF, ClissoldL, WellsR, HattoriC, WernerP, BancroftI. Associative transcriptomics of traits in the polyploid crop species Brassica napus. Nat Biotechnol, 2012, 30: 798-802[本文引用:1][JCR: 32.438]
[22]
CockramJ, WhiteJ, Zuluaga DL, SmithD, ComadranJ, MacaulayM, LuoZ, Kearsey MJ, WernerP, HarrapD, TapsellC, LiuH, Hedley PE, SteinN, SchulteD, SteuernagelB, Marshall DF, Thomas WT, RamsayL, MackayI, Balding DJ, AgouebC, WaughR, O’Sullivan D M. Genome-wide association mapping to cand idate polymorphism resolution in the unsequenced barley genome. Proc Natl Acad Sci USA, 2010, 107: 21611-21616[本文引用:1][JCR: 9.737]
Yan JB, Yang XH, ShahT, Sánchez-VilledaH, Li JS, WarburtonM, ZhouY, Xu YB. High-throughput SNP genotyping with the GoldenGate assay in maize. Mol Breed, 2010, 25: 441-451[本文引用:1][JCR: 3.251]
[27]
Pritchard JK, StephensM, DonnellyP. Inference of population structure using multilocus genotype data. Genetics, 2000, 155: 945-959[本文引用:1][JCR: 4.389]
[28]
EvannoG, RegnautS, GoudetJ. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol, 2005, 14: 2611-2620[本文引用:1][JCR: 6.275]
[29]
ChalhoubB, DenoeudF, LiuS, Parkin IA, TangH, WangX, ChiquetJ, BelcramH, TongC, SamansB, CorréaM, Da SilvaC, JustJ, FalentinC, KohCS, Le ClaincheI, BernardM, BentoP, NoelB, LabadieK, AlbertiA, CharlesM, ArnaudD, GuoH, DaviaudC, AlameryS, JabbariK, ZhaoM, Edger PP, ChelaifaH, TackD, LassalleG, MestiriI, SchnelN, Le Paslier M C, Fan G, Renault V, Bayer P E, Golicz A A, Manoli S, Lee T H, Thi V H, Chalabi S, Hu Q, Fan C, Tollenaere R, Lu Y, Battail C, Shen J, Sidebottom C H, Wang X, Canaguier A, Chauveau A, Bérard A, Deniot G, Guan M, Liu Z, Sun F, Lim Y P, Lyons E, Town C D, Bancroft I, Wang X, Meng J, Ma J, Pires J C, King G J, Brunel D, Delourme R, Renard M, Aury J M, Adams K L, Batley J, Snowdon R J, Tost J, Edwards D, Zhou Y, Hua W, Sharpe A G, Paterson A H, Guan C, Wincker P. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science, 2014, 345: 950-953[本文引用:2]
[30]
LibaultM, WanJ, CzechowskiT, UdvardiM, StaceyG. Identification of 118 Arabidopsis transcription factor and 30 ubiquitin- ligase genes responding to chitin, a plant-defense elicitor. Mol Plant Microbe Interact, 2007, 20: 900-911[本文引用:1][JCR: 4.307]
[31]
Kim YC, NakajimaM, YamaguchiI. Contribution of gibberellins to the formation of Arabidopsis seed coat through starch degradation. Plant Cell Physiol, 2005, 46: 1317-1325[本文引用:1][JCR: 4.134]
[32]
Hunter CA, Aukerman MJ, SunH, FokinaM, Poethig RS. PAUSED encodes the Arabidopsis exportin-t ortholog. Plant Physiol, 2003, 132: 2135-2143[本文引用:1][JCR: 6.555]
[33]
谢田田, 陈玉波, 黄吉祥, 张尧锋, 徐爱遐, 陈飞, 倪西源, 赵坚义. 甘蓝型油菜不同发育时期株高QTL的动态分析. 作物学报, 2012, 38: 1802-1809Xie TT, Chen YB, Huang JX, Zhang YF, Xu AX, ChenF, Ni XY, Zhao JY. Dynamic analysis of QTL for plant height of rapeseed at different developmental stages. Acta Agron Sin, 2012, 38: 1802-1809 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[34]
HunagN, Angeeles ER, DomingoJ. Pyramiding of bacterial blight resistance genes in rice: marker-assisted selection using RFLP and PCR. Theor Appl Genet, 1997, 95: 313-320[本文引用:1][JCR: 3.658]
 , 刘胜毅
, 刘胜毅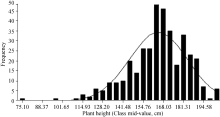




{kind=link}
{kind=link}
{kind=link}