关键词:花生; 休眠性; ABA; AhNCED2; AhCYP707A1 Expression Analysis of Genes Involved in Peanut Seed Dormancy Release ( Arachis hypogaeaL.) CHEN Jing1,4, JIANG Ling1, WANG Chun-Ming1, HU Xiao-Hui4, ZHAI Hu-Qu3, WAN Jian-Min1,2,* 1State Key Laboratory for Crop Genetics and Germplasm Enhancement / Jiangsu Provincial Center of Plant Gene Engineering, Nanjing Agricultural University, Nanjing 210095, China
2Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
3Chinese Academy of Agricultural Sciences, Beijing 100081, China
4 Shandong Peanut Research Institute, Qingdao 266100, China
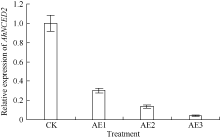
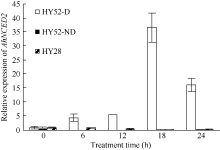
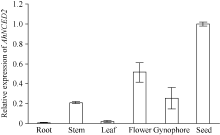
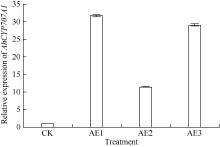
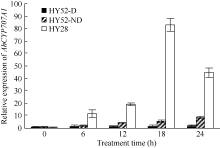
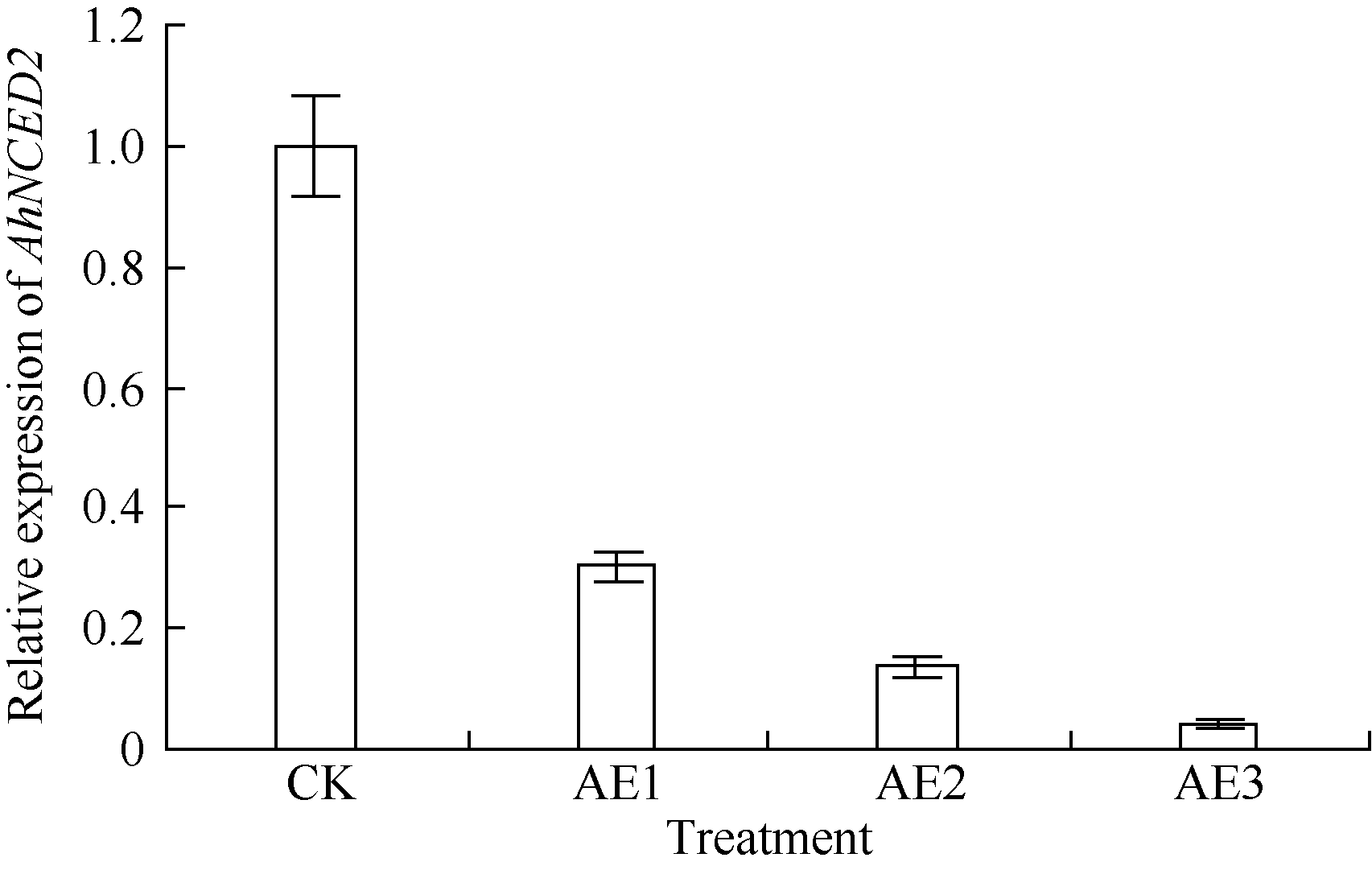
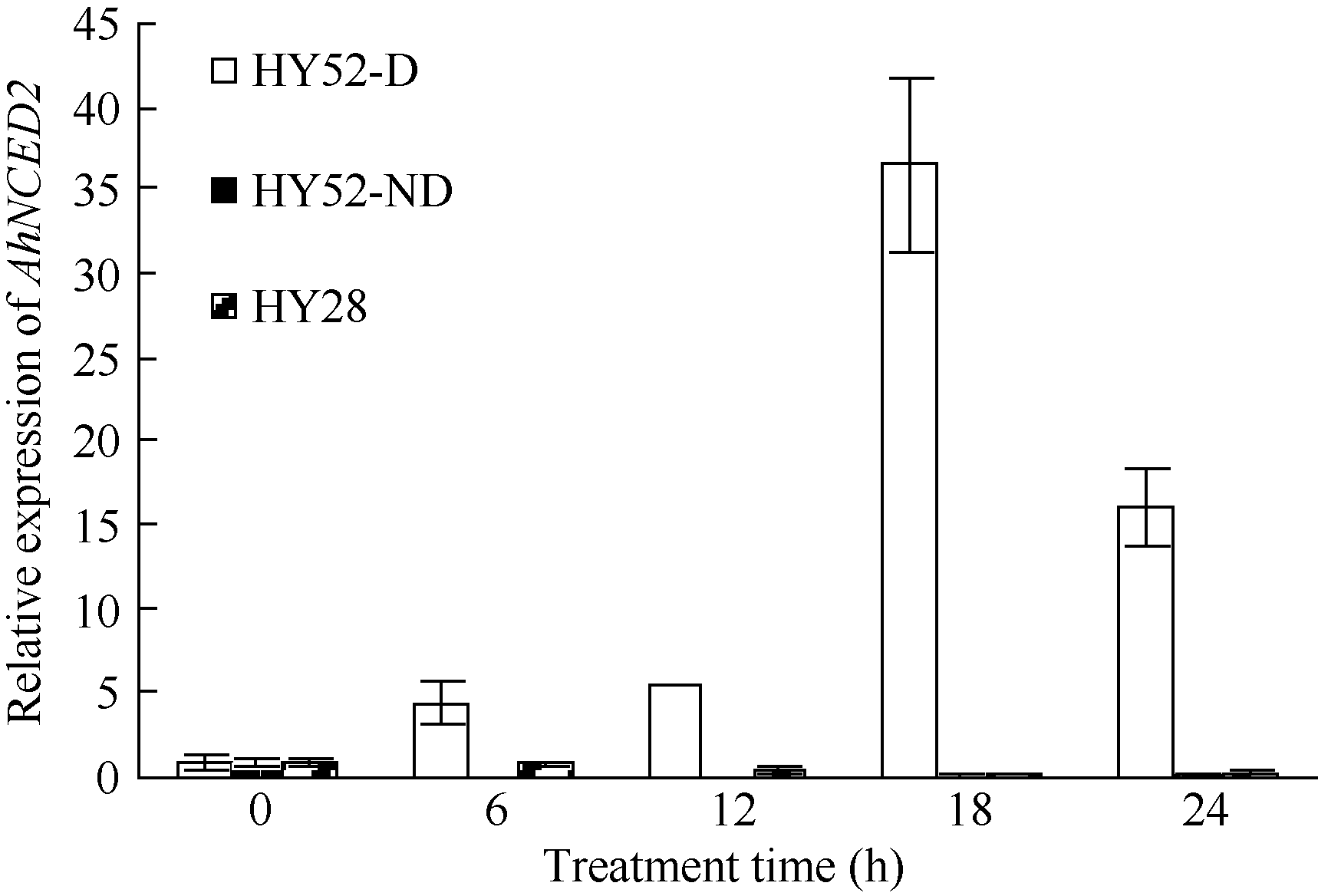
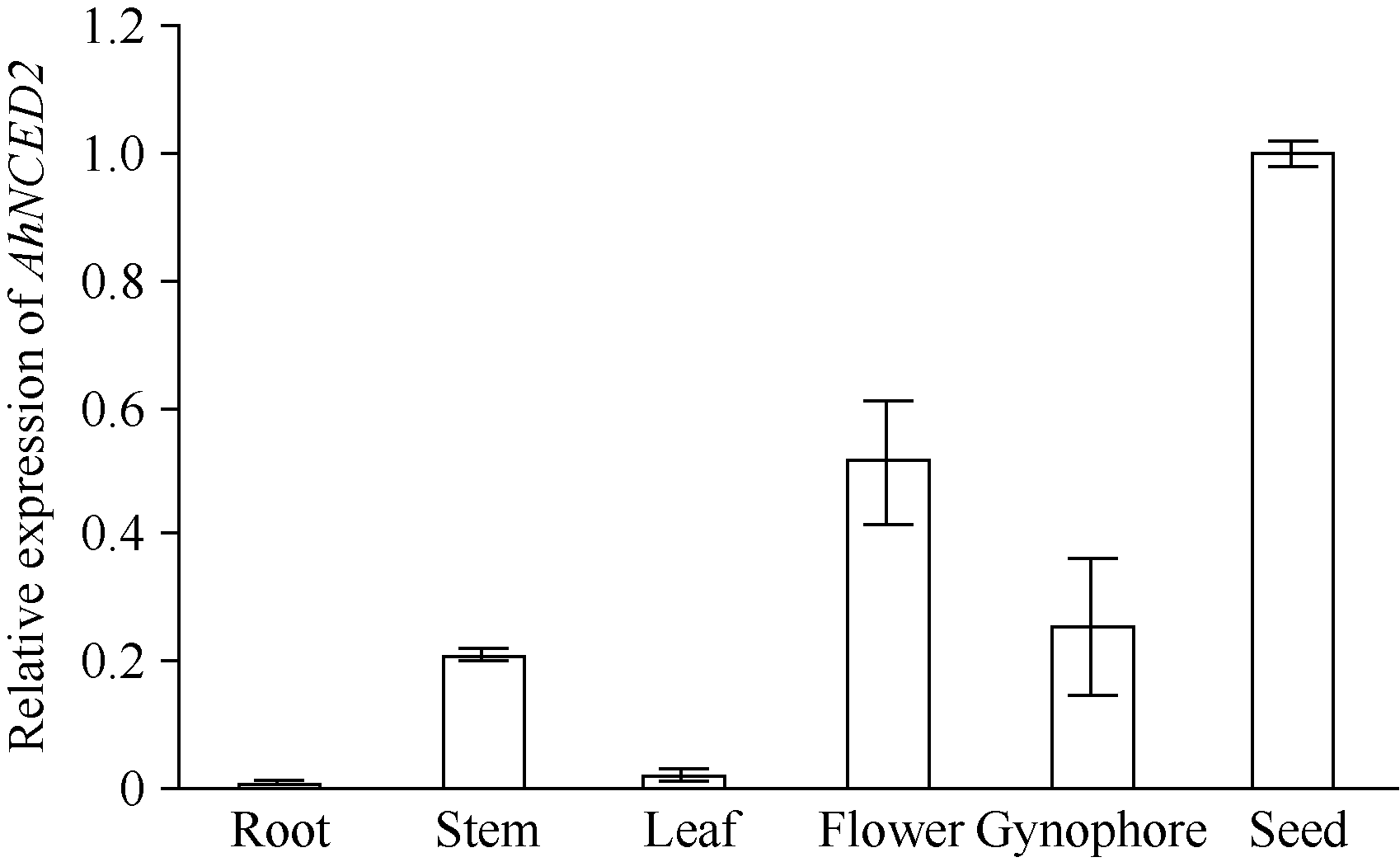
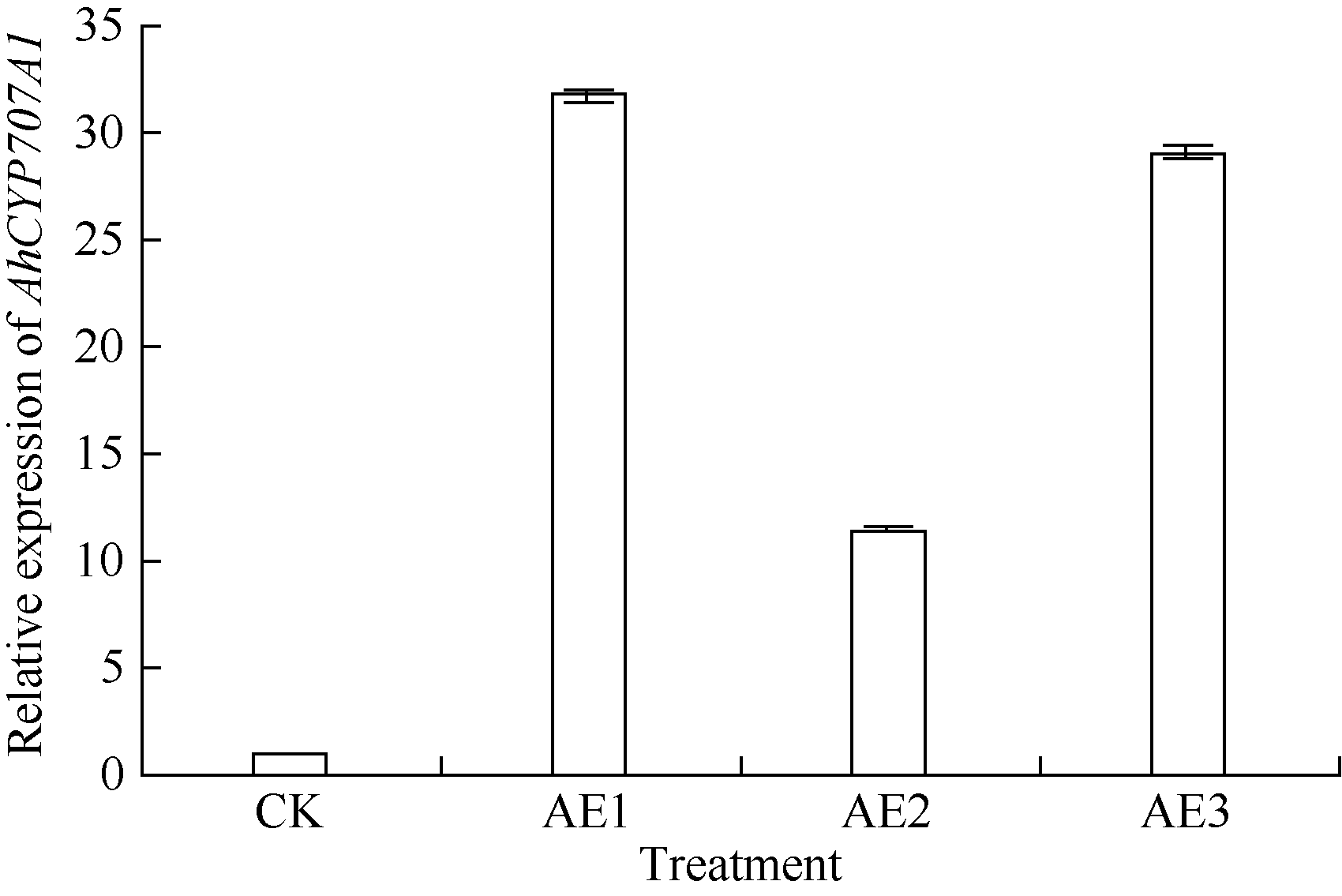
AbstractSeed dormancy is one of important agronomic traits in peanut ( Arachis hypogaea L.). Seed dormancy can be released with exogenous ethephon. To understand the molecular mechanisms of switches from dormancy to germination in peanut seeds underlying the role of ethephon, we preformed transcriptome analyses among imbibed dormant seeds as control and dormancy-released seeds (AE1, AE2, AE3) treated by 100 mg L-1exogenous ethephon, and compared the expression of unigenes related to ABA, GA, ETH and auxin. The results showed that there were 15, 40, 60, and 56 unigenes associated with GA, ABA, ETH, and auxin respectively, which were significantly differentially expressed unigenes during the process from dormancy to germination. The results of Real-time RT-PCR showed that the expressions of AhNCED2and AhCYP707A1 were induced distinctly by exogenous ethephon in seed dormancy released process. In dormant and non-dormant seed imbibition and germination processes, there were different roles between expresses of AhNCED2 and AhCYP707A1. AhNCED2played a positive role in maintaining seed dormancy, while AhCYP707A1played a positive role for seed dormancy breaking.
附表1 种子休眠解除过程中与ABA、GA、ETH、auxin相关的显著性差异表达unigenes Supplementary table 1 Significant differential expression unigenes associated with GA, ABA, ETH, and auxin during the process from dormancy to germination
附表1 种子休眠解除过程中与ABA、GA、ETH、auxin相关的显著性差异表达unigenes Supplementary table 1 Significant differential expression unigenes associated with GA, ABA, ETH, and auxin during the process from dormancy to germination
1.2 文库构建与测序采用Illumina/Solexa标准操作步骤(Directional mRNA-Seq Sample Preparation Part # 15018460 Rev. A, October 2010)制备转录组测序文库。其后用Agilent2100对文库建库片段大小进行质控, 采用 Illumina 2000平台对文库进行测序。 1.3 序列分析和注释对所产生的原始reads进行质量评估和可信度分析, 去除低质量片段(Q< 20)。使用软件Trinity (trinityrnaseq_r2012-10-05)将花生4个样本的有效reads合并进行de novo拼接, 获得121412个Unigene。将获得的Unigenes进行注释, 包括NR、Swiss-Prot等基本数据库注释、KOG分类、GO和KEGG注释。 1.4 差异表达分析使用RPKM (Reads per kb per million reads)计算基因表达量; 根据unigenes表达量采用fold change分析、fisher检验、chisq检验等进行差异表达分析。样本AE1、AE2、AE3均与CK相比较, 取值P≤ 0.05且|fold change|≥ 2。 1.5 花生总RNA的提取及cDNA的合成按照RNA试剂盒(TransGen)说明书提取花生种子总RNA, 用Prime RT Reagent Kit (TaKaRa, 大连)将提取的总RNA反转录成cDNA。 1.6 荧光定量PCR采用LightCycle 2.0 (rRoche Diagnostics公司)荧光定量PCR仪。PCR程序为95℃ 30 s; 95℃ 5 s, 60℃ 20 s, 72℃ 10 s , 45个循环; 然后绘制溶解曲线。采用SYBR Premix Ex Taq试剂盒(TaKaRa, 大连), 按照说明进行实时定量PCR。每个样品重复3次, 取平均值, 采用2-Δ Δ Ct的方法。根据目的基因片段序列, 利用Beacon Designer 7.91软件设计荧光定量PCR引物, 内参基因为Actin11(表2)。 1.7 目的基因片段生物信息学分析利用DNA MAN 6.0对目的基因片段的氨基酸序列、开放阅读框搜索进行分析; 利用在线Blast (http://www.ncbi.nlm.nih.gov/Blast/)进行序列比对、同源序列搜索。 表2 Table 2 表2(Table 2)
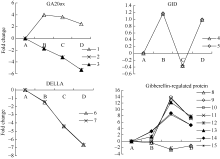
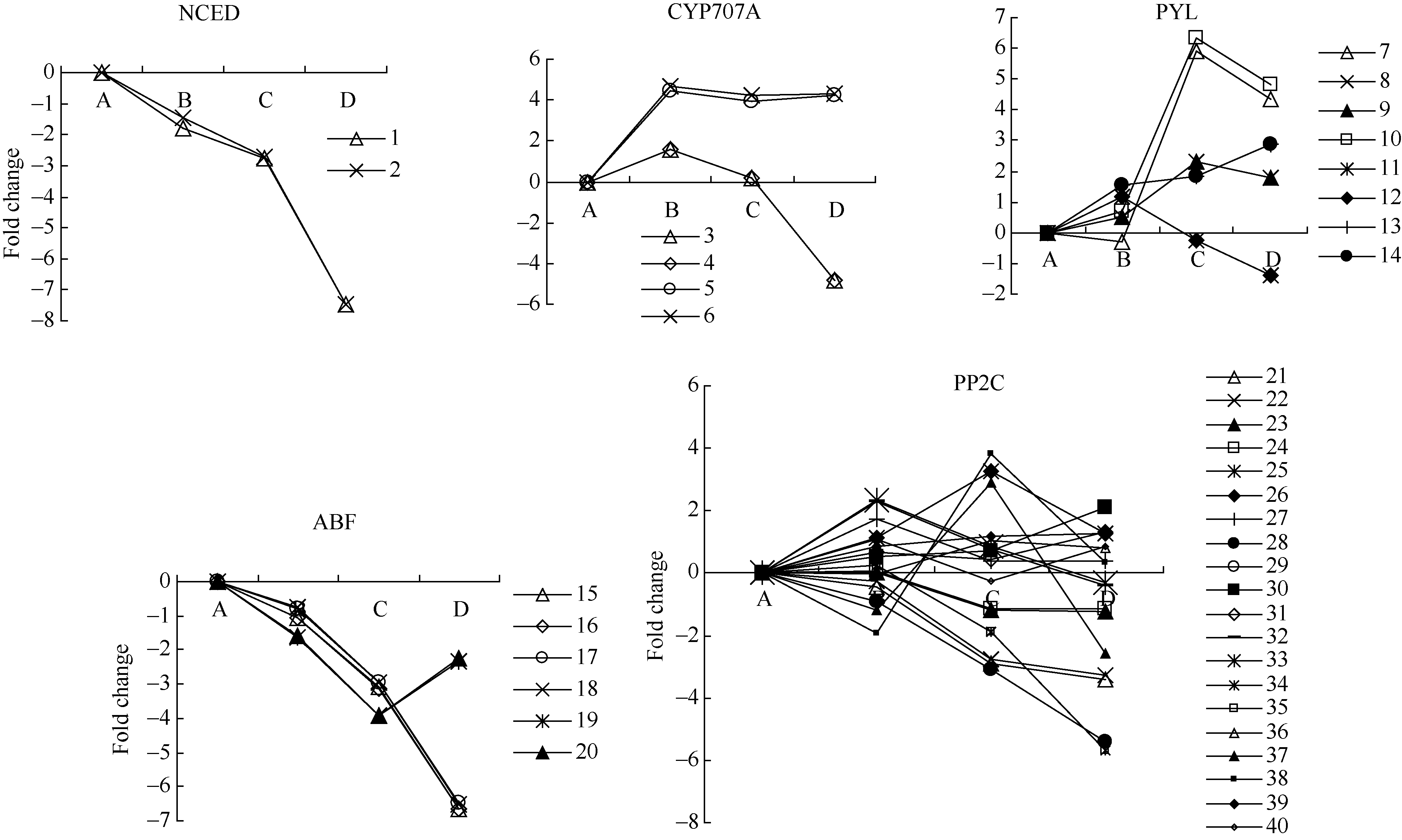
图1 ABA相关unigenes在种子休眠解除过程中的表达变化数字对应unigenes见附表1。 A: log2(CK_RPKM/CK_RPKM); B: log2(AE1_RPKM/CK_RPKM); C: log2(AE2_RPKM/CK_RPKM); D: log2(AE3_RPKM/CK_RPKM). Number corresponding to unigenes see Supplementary table 1.Fig. 1 Expression trend of unigenes related to ABA during the process of seed dormancy release
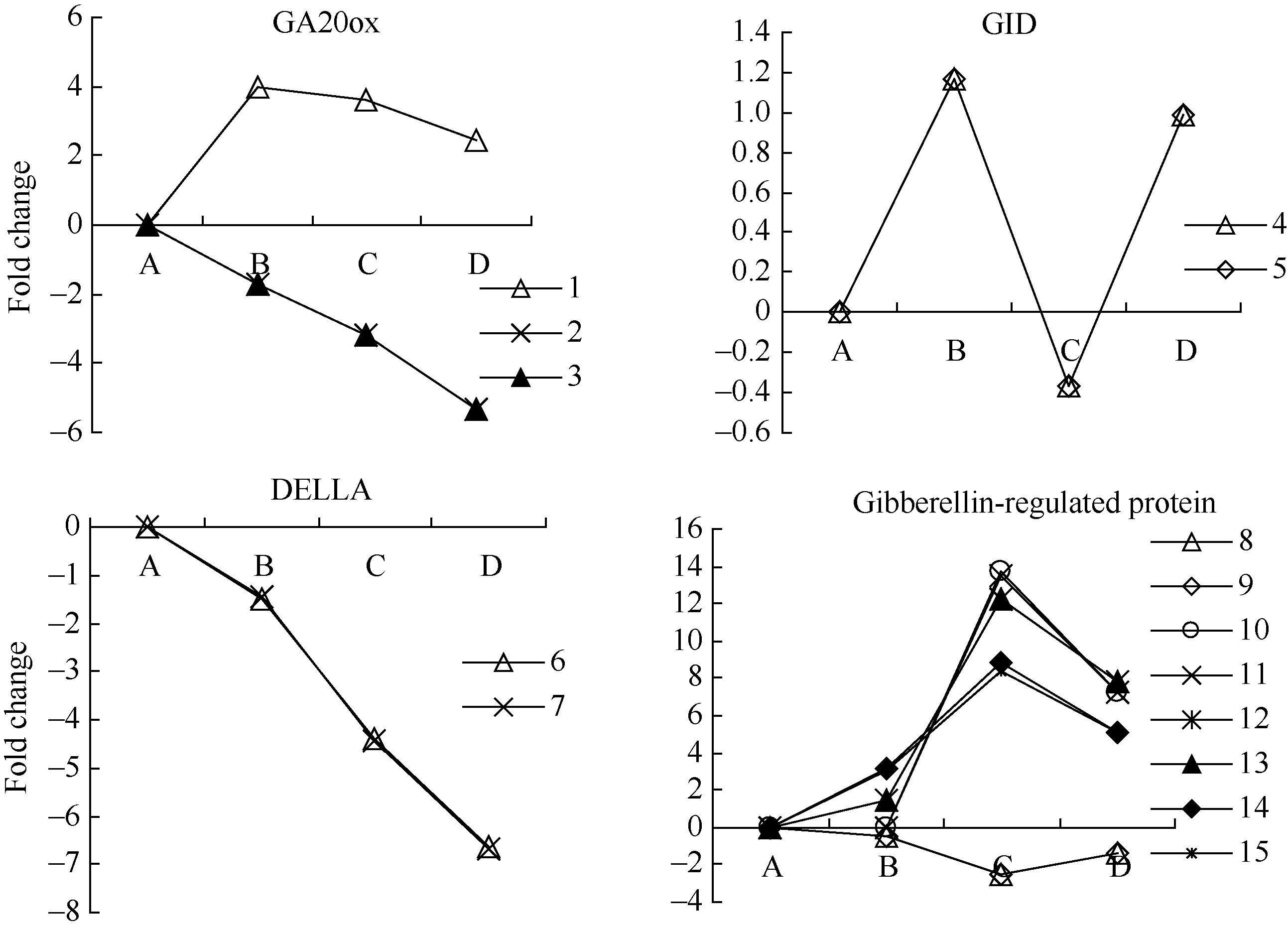
图2 GA相关unigenes在种子休眠解除过程中的表达变化数字对应unigenes见附表1。Fig. 2 Expression trend of unigenes related to GA during the process of seed dormancy releaseA: log2(CK_RPKM/CK_RPKM); B: log2(AE1_RPKM/CK_RPKM); C: log2(AE2_RPKM/CK_RPKM); D: log2(AE3_RPKM/CK_RPKM). Number corresponding to unigenes see Supplementary table 1.
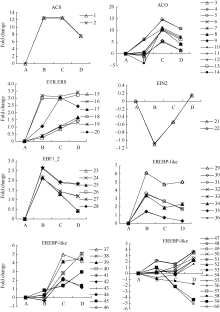
图3 ETH相关unigenes在种子休眠解除过程中的表达变化数字对应unigenes见附表1。Fig. 3 Expression trend of unigenes related to ETH during the process of seed dormancy releaseA: log2(CK_RPKM/CK_RPKM); B: log2(AE1_RPKM/CK_RPKM); C: log2(AE2_RPKM/CK_RPKM); D: log2(AE3_RPKM/CK_RPKM). Number corresponding to unigenes see Supplementary table 1.
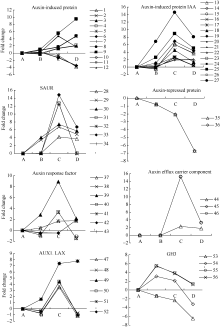
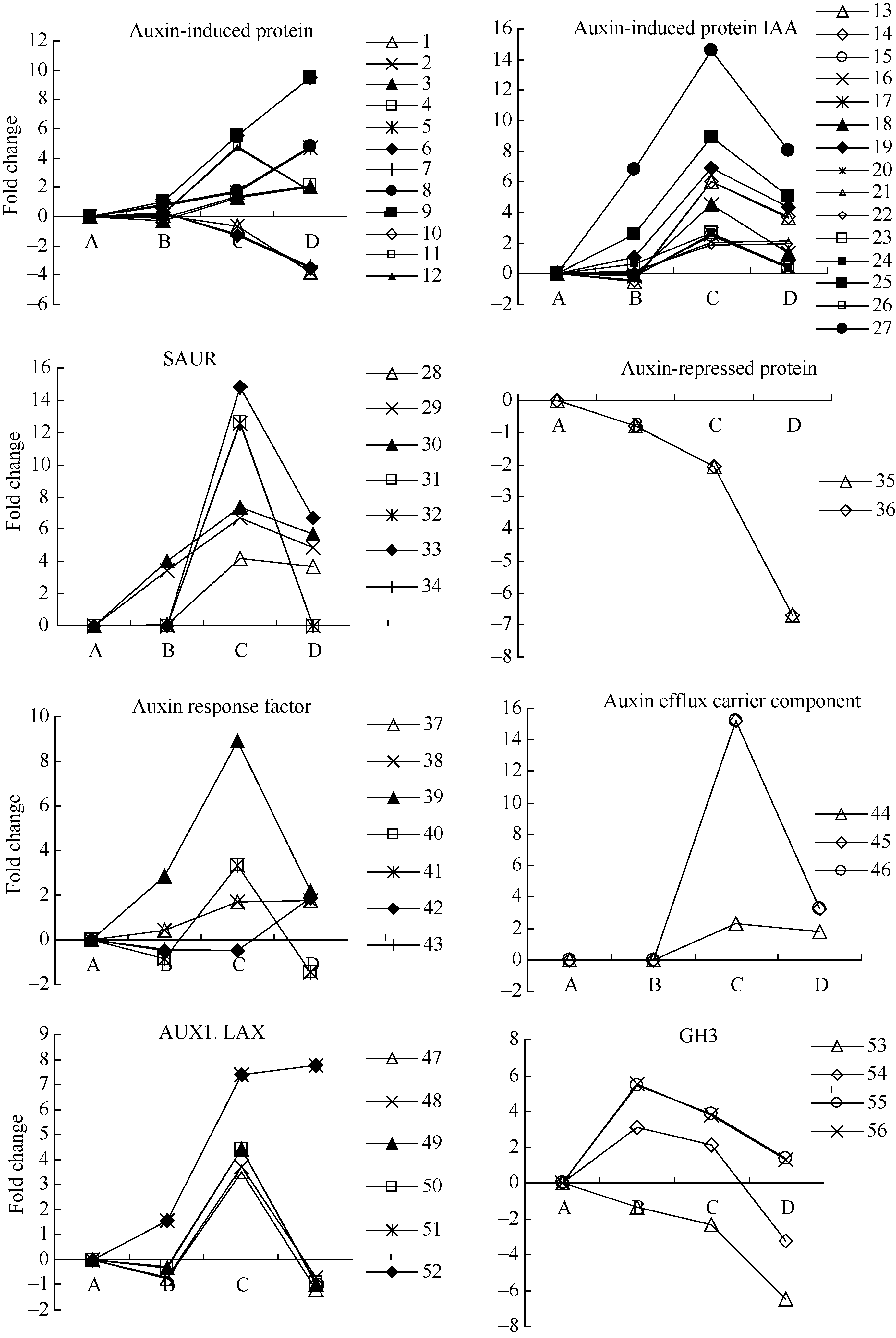
图4 auxin相关unigenes在种子休眠解除过程中的表达变化数字对应unigenes见附表1。Fig. 4 Expression trend of unigenes related to auxin during the process of seed dormancy releaseA: log2(CK_RPKM/CK_RPKM); B: log2(AE1_RPKM/CK_RPKM); C: log2(AE2_RPKM/CK_RPKM); D: log2(AE3_RPKM/CK_RPKM). Number corresponding to unigenes see Supplementary table 1.
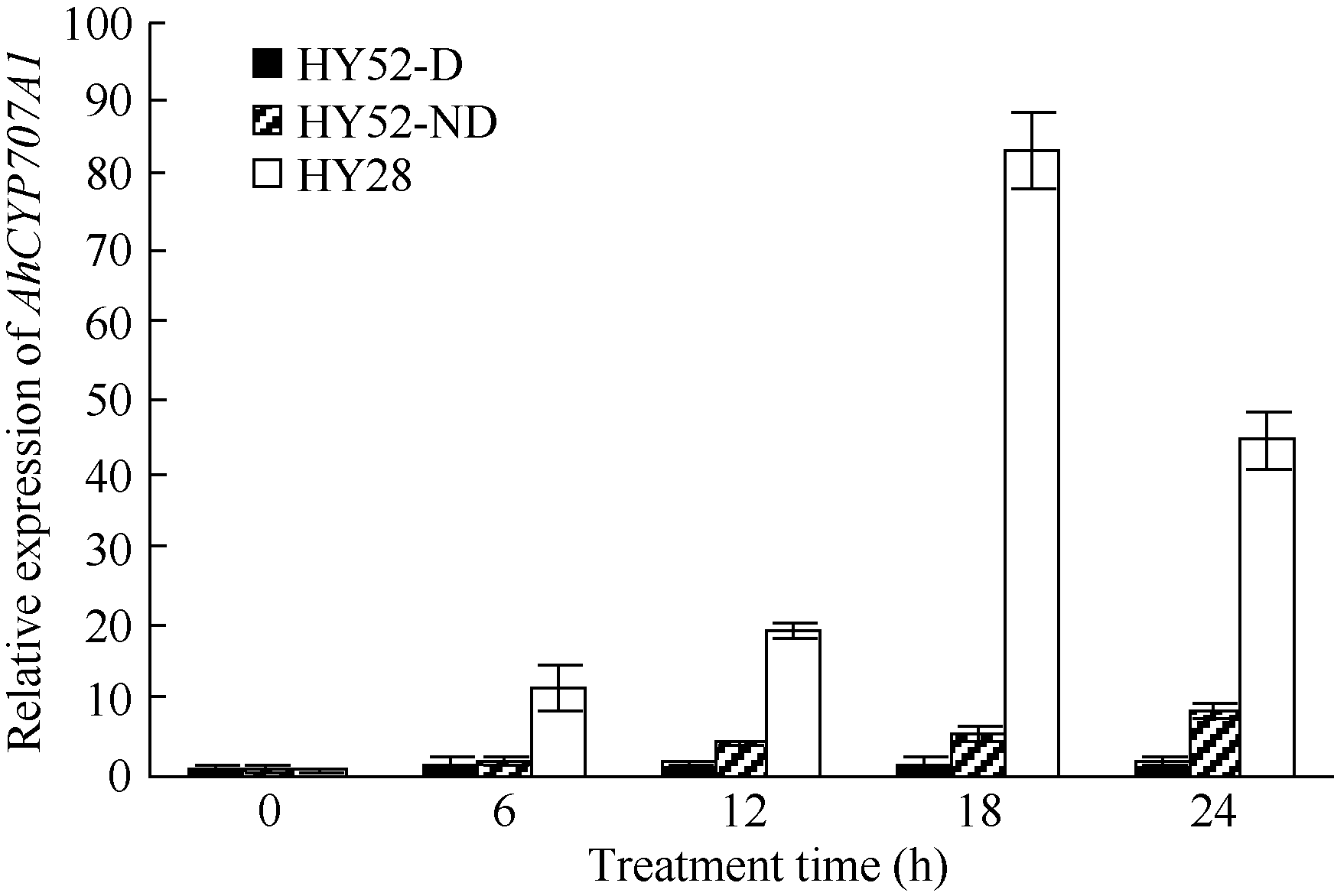
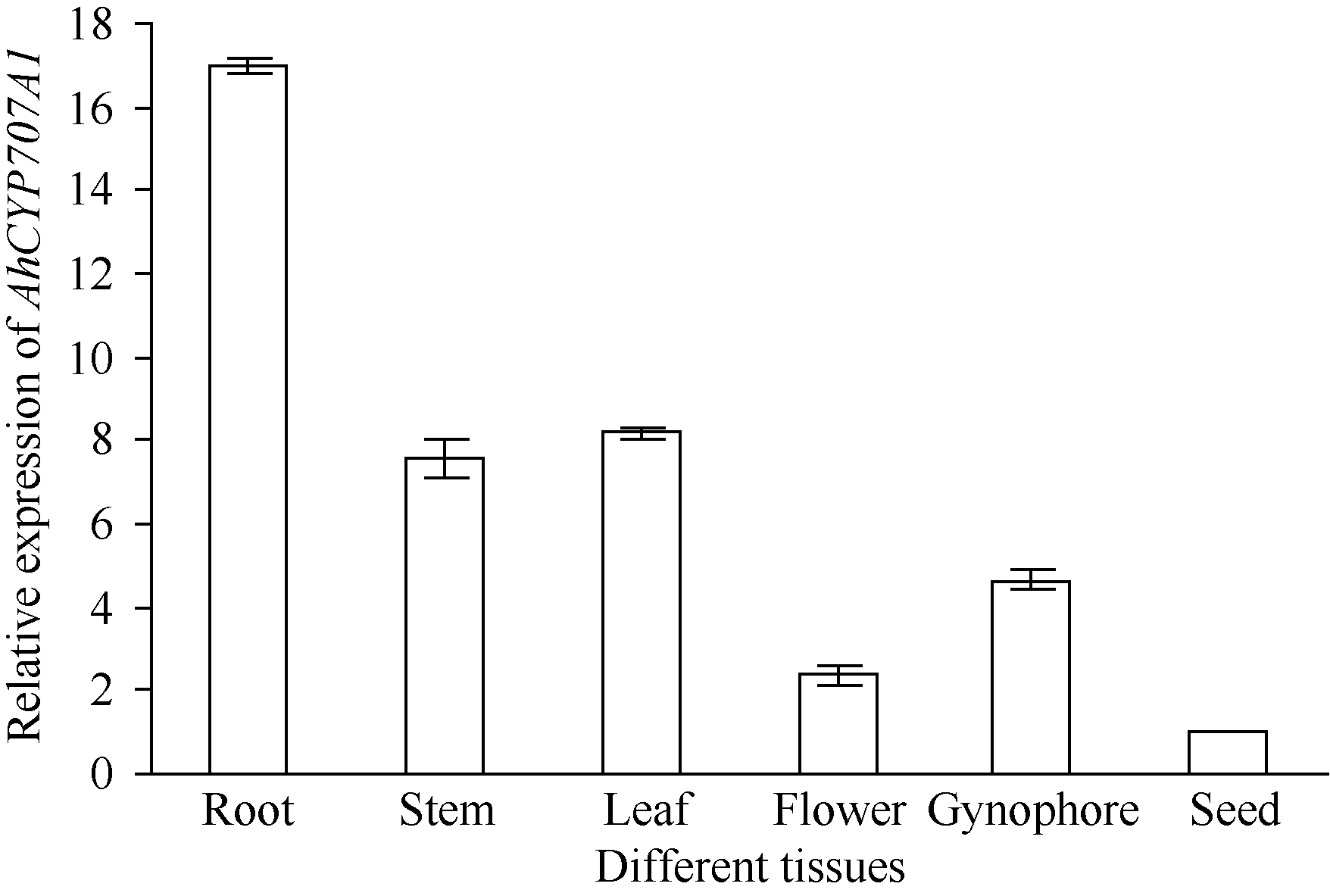
4 结论外源乙烯利诱导花生种子休眠解除过程中, 与GA、ABA、ETH及auxin相关unigenes表现显著差异表达, 表明外源乙烯利通过GA、ABA、ETH、auxin相关基因的诱导作用来完成其对花生种子休眠的解除。AhNCED2和AhCYP707A1受外源乙烯利的诱导, 与花生种子休眠的维持及解除密切相关, 为培育具有适度休眠性的花生新种质提供了候选基因。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
Bewley JD, Bradford KJ, Hilhorst H W M, Nonogaki H. Seed Physiology of Development, Germination and Dormancy (3rd edn). London: Springer, 2013, pp 247-295[本文引用:1]
[2]
曹雅君, 江玲, 罗林广, 翟虎渠, 志村英二, 杨世湖, 万建民. 水稻品种休眠特性的研究. 南京农业大学学报, 2001, 24(2): 1-5Cao YJ, JiangL, LuoLG, Zhai HQ, ShimuraE, Yang SH, Wan JM. A study on seed dormancy in rice (Oryza sativa L. ). J Nanjing Agric Univ, 2001, 24(2): 1-5 (in Chinese with English abstract)[本文引用:1][CJCR: 0.916]
[3]
徐恒恒, 黎妮, 刘树君, 王伟青, 王伟平, 张红, 程红焱, 宋松泉. 种子萌发及其调控的研究进展. 作物学报, 2014, 40: 1141-1156Xu HH, LiN, Liu SJ, Wang WQ, Wang WP, ZhangH, Cheng HY, Song SQ. Research progress in seed germination and its control. Acta Agron Sin, 2014, 40: 1141-1156 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[4]
ErwannA, JulienS, FrancoiseC, LoicR, Marion-poll A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Plant Cell Biol, 2013, 4: 1-19[本文引用:1]
[5]
Ali-RachediS, BouinotD, Wagner MH, BonnetM, SottaB, GrappinP, JullienM. Changes in endogenous abscisic acid levels during dormancy release and maintenance of mature seeds: studies with the Cape Verde Island s ecotype, the dormant model of Arabidopsis thaliana. Planta, 2004, 219: 479-488[本文引用:1][JCR: 3.347]
[6]
Cadman CS, Toorop PE, Hilhorst HW, Finch-Savage W E. Gene expression profiles of Arabidopsis Cvi seeds during dormancy cycling indicate a common underlying dormancy control mechanism. Plant J, 2006, 46: 805-822[本文引用:1][JCR: 6.582]
[7]
NambaraE, OkamotoM, TatematsuK, YanoR, SeoM, KamiyaY. Abscisic acid and the control of seed dormancy and germination. Seed Sci Res, 2010, 20: 55-67[本文引用:1][JCR: 1.931]
[8]
MatakiadisT, AlboresiA, JikumaruY, TatematsuK, PichonO, Renou JP, Kamiya YJ, NambarE, Truong HN. The Arabidopsis abscisic acid catabolic gene CYP707A2 plays a key role in nitrate control of seed dormancy. Plant Physiol, 2009, 149: 949-960[本文引用:2][JCR: 6.555]
[9]
OkamotoM, KuwaharaA, SeoM, KushiroT, AsamiT, HiraiN, KamiyaY, KoshibaT, NambaraE. CYP707A1 and CYP707A2, which encode abscisic acid 8’-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiol, 2006, 141: 97-107[本文引用:2][JCR: 6.555]
[10]
PenfieldS, LiY, Gilday AD, GrahamS, Graham IA. Arabidopsis ABA INSENSITIVE4 regulates lipid mobilization in the embryo and reveals repression of seed germination by the endosperm. Plant Cell, 2006, 18: 1887-1899[本文引用:1][JCR: 9.251]
[11]
Matilla AJ. Ethylene in seed formation and germination. Seed Sci Res, 2000, 10: 111-126[本文引用:1][JCR: 1.931]
[12]
SiriwitayawanG, Geneve RL, Downie AB. Seed germination of ethylene perception mutants of tomato and Arabidopsis. Seed Sci Res, 2003, 13: 303-314[本文引用:2][JCR: 1.931]
[13]
Chiwocha S DS, Cutler AJ, Abrams SR, Ambrose SJ, YangJ, Ross A RS, Kermode A R The etr1-2 mutation in Arabidopsis thaliana affects the abscisic acid, auxin, cytokinin and gibberellin metabolic pathways during maintenance of seed dormancy, moist-chilling and germination. Plant J, 2005, 42: 35-48[本文引用:1][JCR: 6.582]
[14]
Karssen CM, ZagorskiS, KepczynskiJ, Groot S P C. Key role for endogenous gibberellins in the control of seed germination. Ann Bot (London), 1989, 63: 71-80[本文引用:2][JCR: 3.449]
[15]
KoornneefM, Karssen CM. Seed dormancy and germination. In: Meyerowitz E M, Sommerville C R, eds. Arabidopsis. New York: Cold Spring Harbor Laboratory, 1994. pp 313-334[本文引用:1]
[16]
禹山林. 中国花生遗传育种学. 上海: 上海科学技术出版社, 2011Yu SL. Genetics and Breeding of China peanut. Shanghai: Shanghai Science and Technology Press, 2011 (in Chinese)[本文引用:1]
[17]
周桂元, 梁炫强. 花生种子休眠性的研究进展. 江西农业学报, 2011, 23(11): 61-63Zhou GY, Liang XQ. Research advance in seed dormancy of peanut. Acta Agric Jiangxi, 2011, 23: 61-63 (in Chinese with English abstract)[本文引用:1][CJCR: 0.9866]
[18]
Kumarl SS, Patel SA. Seed dormancy in groundnut (Arachis hypogaea L. ): II. Estimation of gene effects in six crosses. Trop Agric (Trinidad), 1999, 76: 188-192[本文引用:1]
[19]
Upadhyaya HD, Nigam SN. Inheritance of fresh seed dormancy in peanut. Crop Sci, 1999, 39: 98-101[本文引用:1][JCR: 1.513]
[20]
胡晓辉, 苗华荣, 杨伟强, 张建成, 陈静. 花生种子休眠性的遗传分析及其影响因素的研究. 核农学报, 2013, 27: 1449-1455Hu XH, Miao HR, Yang WQ, Zhang JC, ChenJ. Genetic analysis and factors influencing peanut (Arachis hypogaea L. ) seed dormancy. J Nucl Agric Sci, 2013, 27: 1449-1455 (in Chinese with English abstract)[本文引用:1][CJCR: 1.237]
Ketring DL, Morgan PW. P hysiology of Oil Seeds I. Regulation of dormancy in Virginia-type peanut seed. Plant Physiol, 1970, 45: 268-273[本文引用:1][JCR: 6.555]
[23]
Ketring DL, Morgan PW. Physiology of oil seeds: II. Dormancy release in Virginia-type peanut seeds by plant growth regulators. Plant Physiol, 1971, 47: 488-492[本文引用:1][JCR: 6.555]
[24]
Ketring DL, Morgan PW. Ethylene as a component of the emanations from germinating peanut seeds and its effect on dormant Virginia-type seeds. Plant Physiol, 1969, 44: 326-330[本文引用:1][JCR: 6.555]
[25]
Wang ML, Chen CY, Pinnow DL, Barkley NA, Pittman RN, LambM, Pederson GC. Seed dormancy variability in the US peanut Mini-core collection. Res J Seed Sci, 2012, 5: 84-95[本文引用:1]
[26]
IssaF, DanièlF, Jean-FrançoisR, Hodo-AboloT, Mbaye-NdoyeS, Tahir DA, OusmaneN. Inheritance of fresh seed dormancy in Spanish-type peanut (Arachis hypogaea L. ): bias introduced by inadvertent selfed flowers as revealed by microsatellite markers control. Afr J Biotechnol, 2010, 9: 1905-1910[本文引用:1][JCR: 0.573]
Gonzalez-GuzmanM, AbiaD, SalinaJ, SerranoR, RodriguezP. Two new alleles of the abscisic aldehyde oxidase 3 gene reveal its role in abscisic acid biosynthesis in seeds. Plant Physiol, 2004, 135: 325-333[本文引用:1][JCR: 6.555]
[32]
KushiroT, OkamotoM, NakabayashiK, YamagishiK, KitamuraS, AsamiT, HiraiN, KoshibaT, KamiyaY, NambaraE. The Arabidopsis cytochrome P450 CYP707A encodes ABA 8’-hydroxylases: key enzymes in ABA catabolism. EMBO J, 2004, 23: 1647-1656[本文引用:1][JCR: 9.822]
[33]
ChonoM, HondaI, ShinodaS, KushiroT, KamiyaY, NambaraE, KawakamiN, KanekoS, WatanabeY. Field studies on the regulation of abscisic acid content and germinability during grain development of barley: molecular and chemical analysis of pre-harvest sprouting. J Exp Bot, 2006, 57: 2421-2434[本文引用:1][JCR: 5.242]
[34]
Millar AA, Jacobsen JV, Ross JJ, Helliwell CA, Poole AT, ScofieldG, Reid JB, GublerF. Seed dormancy and ABA metabolism in Arabidopsis and barley: the role of ABA 8’-hydroxylase. Plant J, 2006, 45: 942-954[本文引用:1][JCR: 6.582]
[35]
Thompson AJ, Jackson AC, Parker RA, Morpeth DR, BurbidgeA, Taylor IB. Abscisic acid biosynthesis in tomato: regulation of zeaxanthin epoxidase and 9-cis-epoxycarotenoid dioxygenase mRNAs by light/dark cyeles, water stress and abscisic acid. Plant Mol Biol, 2000, 42: 833-845[本文引用:1][JCR: 3.518]
[36]
White CN, Proebsting WM, HeddenP, White CN, Rivin CJ. Gibberellins and seed development in maize: I. Evidence that gibberelllin/abscisic acid balance governs germination versus maturation pathways. Plant Physiol, 2000, 122: 1081-1088[本文引用:1][JCR: 6.555]
[37]
Calvo AP, Nicola SC, NicolasG, RodriguezD. Evidence of a cross-talk regulation of a GA 20-oxidase (FsGA20ox1) by gibberellins and ethylene during the breaking of dormancy in Fagus sylvatica seeds. Physiol Plant, 2004, 120: 623-6301[本文引用:1][JCR: 6.555]
[38]
HermannK, MeinhardJ, DobrewP, LinkiesA, PesekB, HessB, MachackovaI, FischerU, Leubner-MetzgerG. 1-aminocyclopropane-1-carboxylic acid and abscisic acid during the germination of sugar beet (Beta vulgaris L. ): a comparative study of fruits and seeds. J Exp Bot, 2007, 58: 3047-3060[本文引用:2][JCR: 5.242]
[39]
Leubner-MetzgerG, PetruzzelliL, WaldvogelR, Vogeli-LangeR, MeinsE. Ethylene-responsive element binding protein (EREBP) expression and the transcriptional regulation of class I beta-1, 3-glucanase during tobacco seed germination. Plant Mol Biol, 1998, 38: 785-795[本文引用:2][JCR: 3.518]
[40]
PetruzzelliL, CoraggioL, Leubner-MetzgerG. Ethylene promotes ethylene biosynthesis during pea seed germination by positive feedback regulation of 1-aminocyclo-propane-1-carboxylic acid oxidase. Planta, 2000, 211: 144-149[本文引用:2][JCR: 3.347]
[41]
LinkiesA, MüllerK, MorrisK, TureckovaV, WenkM, Cadman C S C, Corbineau F, Stmad M, Lynn J R, Finch-Savage W E, Leubner-Metzger G. Ethylene interacts with abscisic acid to regulate endosperm rupture during germination: a comparative approach using Lepidium sativum and Arabidopsis thaliana. Plant Cell, 2009, 21: 3803-3822[本文引用:2]
[42]
PetruzzelliL, SturaroM, MainieriD, Leubner-MetzgerG. Calcium requirement for ethylene-dependent responses involving 1-aminocyclopropane-1-carboxylic acid oxidase in radical tissues of germinated pea seeds. Plant Cell Environ, 2003, 26: 661-671[本文引用:1][JCR: 5.135]
[43]
Teale WD, Paponov IA, PalmeK. Auxin in action: signalling transport and the control of plant growth and development. Nat Rev Mol Cell Biol, 2006, 7: 847-859[本文引用:1]
[44]
LeyserO. Dynamic integration of auxin transport and signalling. Curr Biol, 2006, 16: 424-433[本文引用:1][JCR: 9.494]
Liu XD, ZhangH, ZhaoY, FengZ, LiQ, Yang HQ, LuanS, LiJ, He ZH. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc Natl Acd Sci USA, 2013, 110: 15485-15490[本文引用:1]
[47]
Matilla AJ, Matilla-Vazquez M A. Involvement of ethylene in seed physiology. Plant Sci, 2008, 175: 87-97[本文引用:1][JCR: 2.922]
[48]
GhassemianM, NambaraE, CutlerS, KawaideH, KamiyaY, McCourt P. Regulation of abscisic acid signaling by the ethylene response pathway in Arabidopsis. Plant Cell, 2000, 12: 1117-1126[本文引用:1][JCR: 9.251]
[49]
KendeH, Van der Knaap E, Cho H T. Deepwater rice: a model plant to study stem elongation. Plant Physiol, 1998, 118: 1105-1110[本文引用:2][JCR: 6.555]
[50]
BeaudoinN, SerizetC, GostiF, GiraudatJ. Interactions between abscisic acid and ethylene signaling cascades. Plant Cell, 2000, 12: 1103-1116[本文引用:1][JCR: 9.251]
GazzarriniS, McCourt P. Genetic interactions between ABA, ethylene and sugar signaling pathways. Curr Opin Plant Biol, 2001, 4: 387-391[本文引用:1][JCR: 8.455]
[53]
KuceraB, Cohn MA, Leubner-MetzgerG. Plant hormone interactions during seed dormancy release and germination. Seed Sci Res, 2005, 15: 281-307[本文引用:1][JCR: 1.931]
[54]
LiuS, LvY, Wan XR, LiLM, HuB, LiL. Cloning and expression analysis of cDNAs encoding ABA 8’-hydroxylase in peanut plants in response to osmotic stress. PLoS One, 2014, 5: e97025[本文引用:1][JCR: 3.73]
 , 江玲
, 江玲








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}