关键词:水稻; 斑点叶突变体; 白叶枯病抗性; 遗传分析; 基因定位 Characterization of a Novel Spotted Leaf Mutant spl32 and Mapping of Spl32(t) Gene in Rice ( Oryza sativa) ZHONG Zhen-Quan**, LUO Wen-Long**, LIU Yong-Zhu, WANG Hui, CHEN Zhi-Qiang*, GUO Tao* National Engineering Research Centre of Plant Space Breeding / South China Agricultural University, Guangzhou 510642, China
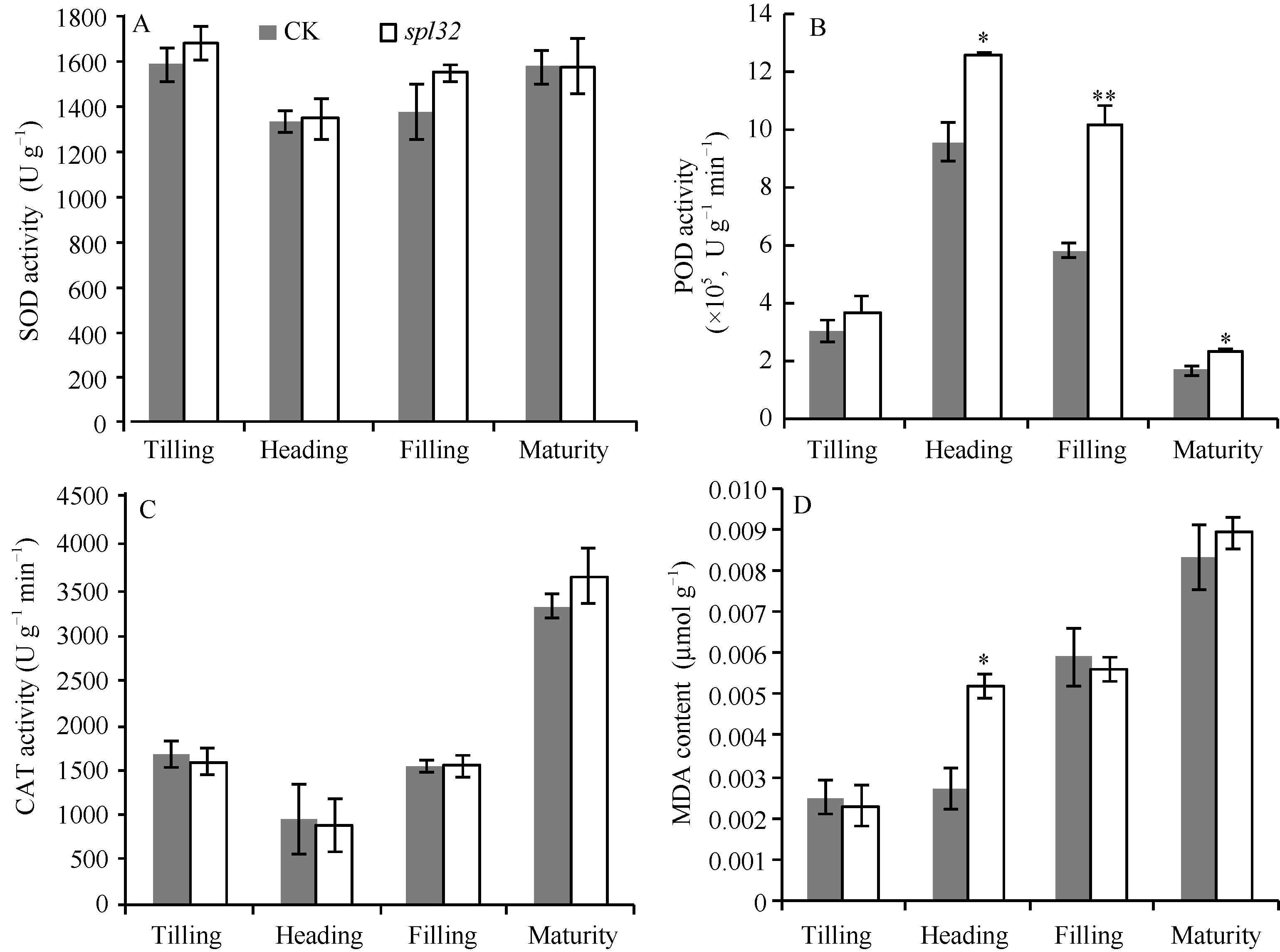
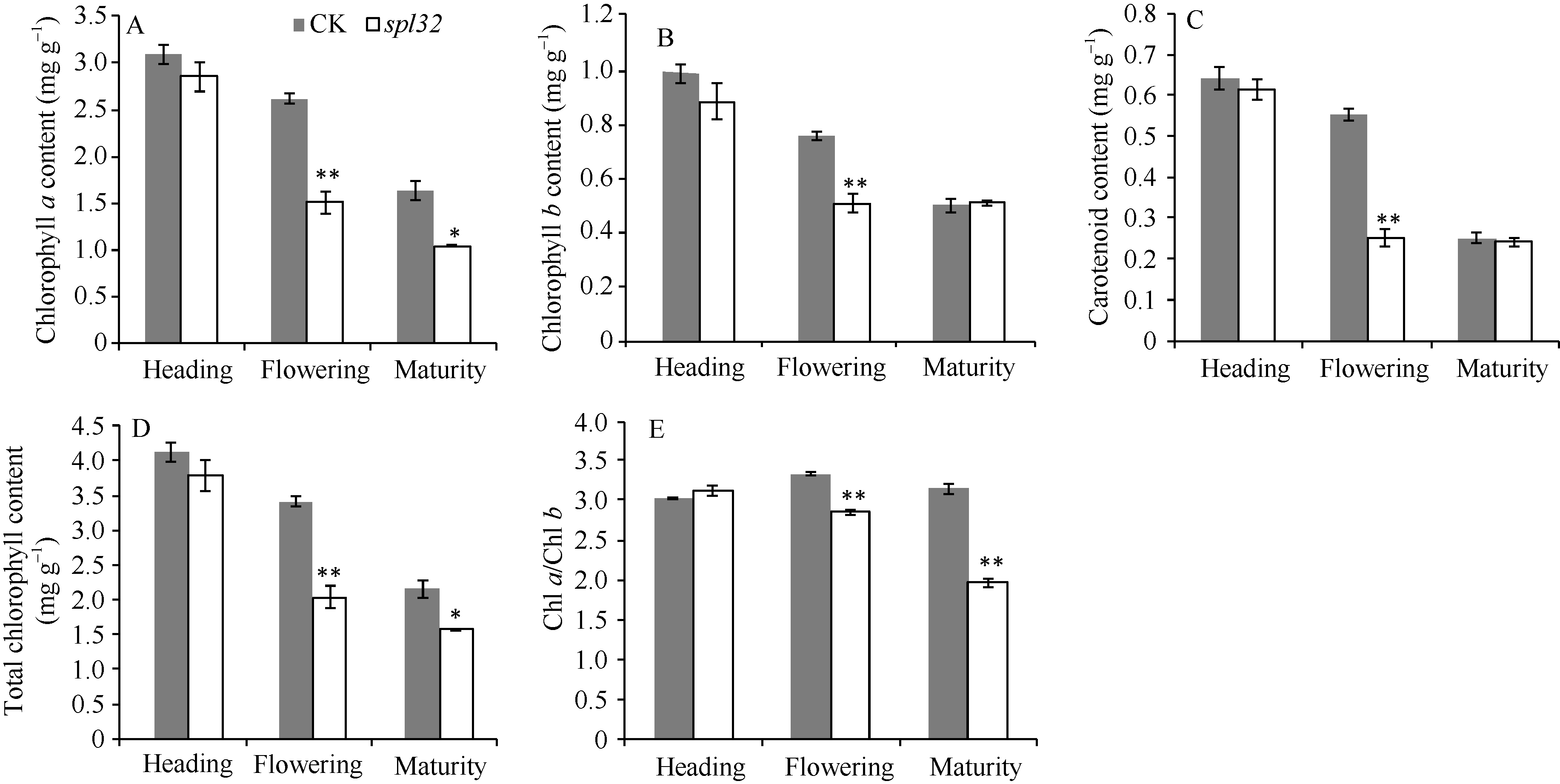
AbstractA dominant spotted-leaf mutant of rice was isolated from F2 (Yuejingsimiao 2/H4) population. The mutant, designated as spl32 ( spotted-leaf 32), initiated brown spots on leaf apex at the panicle differentiation period, and then gradually spread them to whole leaf and sheath. Trypan blue staining indicated that the formation of spots was not caused by cell death. Taken normal green leaf plants segregated from heterozygous F5 as control (CK), we found seeds per panicle and seed setting rate of spotted leaf plants were significantly lower than these of CK. After appearance of spots, the POD activity and MDA content of spl32 were significantly higher than these of CK, while photosynthetic pigment content in spl32was reduced, without significant changes in chlorophyll fluorescence parameters. The resistance to rice bacterial blight in spl32was greatly improved by inoculation of Xanthomonas oryzae pv. oryzae at heading period. The spotted-leaf trait of spl32was verified to be controlled by a dominant gene that temporarily designated as Spl32(t). The novel rice spotted-leaf gene was mapped between markers Ind-c and RM206 on chromosome 11 with a F2 (02428/ spl32) population.
Keyword:Rice ( Oryza sativaL.); Spotted-leaf mutants; Bacterial blight resistance; Genetic analysis; Mapping Show Figures Show Figures
图1 斑点叶突变体spl32的表型A: 灌浆期spl32的田间表型; B: 对照(左)和spl32 (右)乳熟期的植株表型; C: 对照(左)和spl32 (右)灌浆期叶片; D: 成熟期spl32的叶鞘; E: 灌浆期spl32的叶片和穗部。Fig. 1 Phenotypes of spl32A: field phenotype of spl32during the filling stage; B: phenotypes of CK (left) and spl32 (right) during the milky stage; C: leaves of CK (left) and spl32 (right) during the filling stage; D: sheath of spl32during the mature stage; E: phenotypes of leaf and panicle during the filling stage in spl32.
表1 Table 1 表1(Table 1)
表1 对照(CK)和spl32的农艺性状分析 Table 1 Agronomic traits of the control (CK) and spl32
材料 Material
株高 Plant height (cm)
单株穗重 Panicle weight per plant (g)
有效穗数 Number of panicles
每穗粒数 Grain number per panicle
每穗实粒数 Filled grain number per panicle
结实率 Seed setting rate (%)
千粒重 1000-grain weight (g)
CK
93.60± 1.69
22.43± 2.83
6.33± 0.21
235.80± 7.22
217.90± 6.73
92.41± 0.48
19.68± 0.29
spl32
95.08± 1.02
20.37± 1.41
7.33± 0.61
214.87± 4.81*
183.20± 5.54* *
85.13± 1.45* *
19.24± 0.15
* Significantly different at P< 0.05; * * significantly different at P< 0.01. * 在0.05水平上差异显著; * * 在0.01水平上显著差异。
表1 对照(CK)和spl32的农艺性状分析 Table 1 Agronomic traits of the control (CK) and spl32
图2 遮光对突变体spl32叶片的影响A: 对照; B: 成熟期spl32叶片; C: 未发生斑点部位遮光7 d后; D: 遮光部位恢复光照7 d后; E: 已有斑点部位遮光7 d后。Fig. 2 Effects of shading on spl32leavesA: CK; B: spl32 during the maturity stage; C: part of non-spotted leaf after shading for seven days; D: leaf shaded for seven days then under normal light for seven days; E: spotted leaf after shading for seven days.
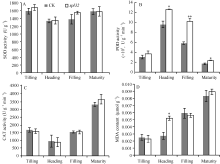
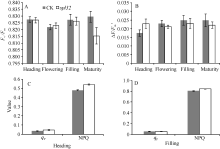
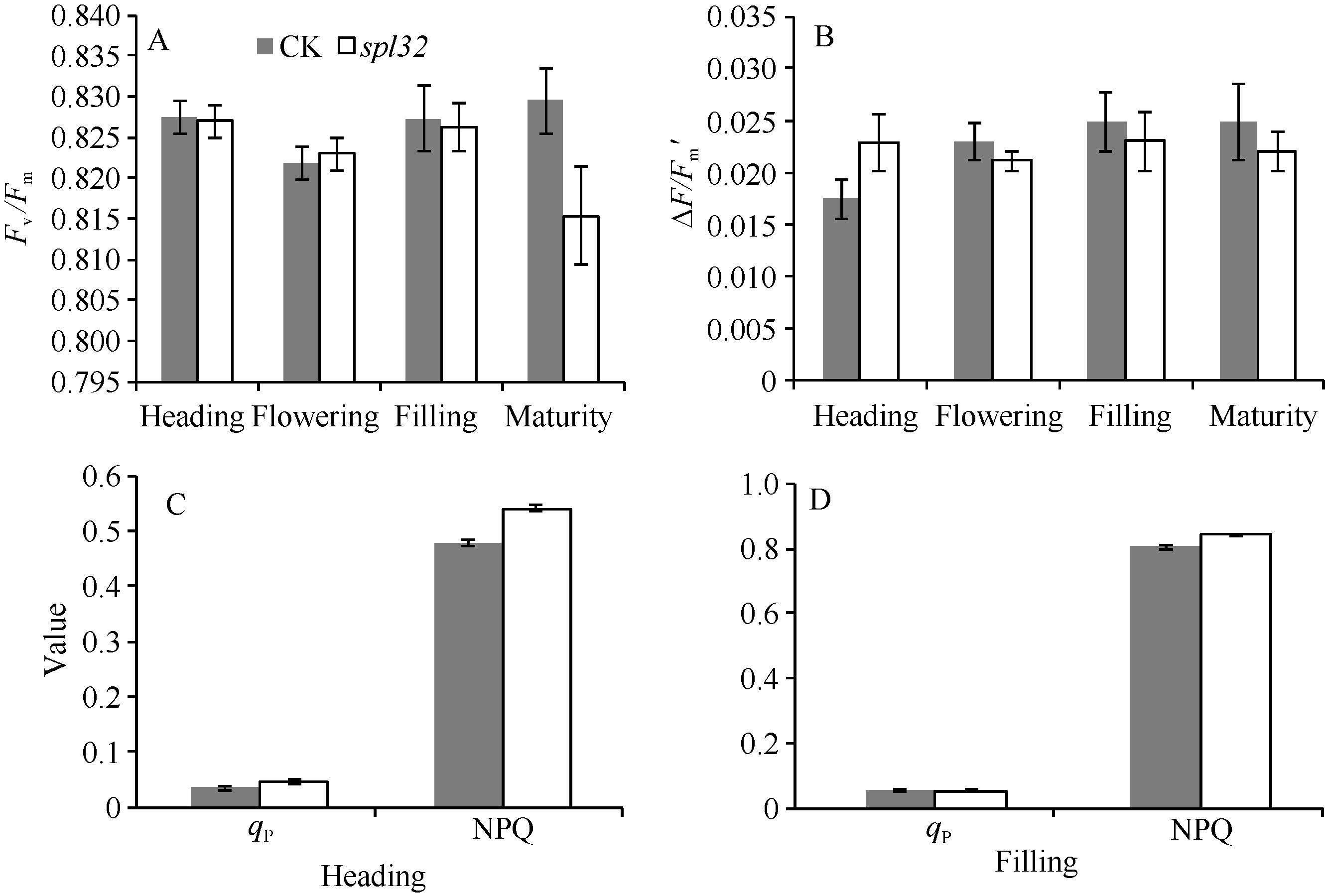
图6 突变体spl32与对照(CK)的叶绿素荧光动力学参数比较Fig. 6 Chlorophyll fluorescence kinetic parameters of the spl32 mutant and control (CK)
A: CK和spl32在各时期叶片的Fv/Fm比较; B: CK和spl32在各时期叶片的∆ F/Fm'比较; C: 抽穗期CK和spl32的qP和NPQ值比较; D: 灌浆期CK和spl32的qP和NPQ值比较。 A: Fv/Fm of spl32 and check during different growth stages; B: ∆ F/Fm' of spl32 and check during different growth stages; C: the qP and NPQ of spl32and check during heading stage; D: the qP and NPQ of spl32 and check during filling stage. 图7 Fig. 7
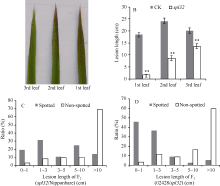
图7 斑点叶性状与白叶枯病抗性的相关性分析A: 接菌时3片功能叶的斑点情况; B: 3片功能叶的病斑调查; C: F2(spl32/日本晴); D: F2(02428/spl32)。Fig. 7 Association analysis of the spotted-leaf trait and bacterial blight resistance
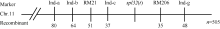
* 在0.05水平上差异显著; * * 在0.01水平上差异显著。 A: spots on the three functional leaves when inoculated; B: lesion length of the three functional leaves in CK and Spl32(cm); C: F2(spl32/Nipponbare); D: F2 (02428/spl32). * Significantly different at P< 0.05; * * Significantly different at P< 0.01. 2.8 Spl32(t)基因的遗传分析与分子定位spl32分别与日本晴和02428配制的F1均表现为斑点叶表型, 说明突变表型由显性基因控制。分别统计F2(02428/spl32)群体和F2(spl32/日本晴)群体中的斑点叶和正常绿叶个体数, 调查其分离比例(表2)表明, 该斑点叶性状受1个显性核基因控制, 暂命名为Spl32(t)。 选用02428× spl32杂交的F2群体为定位群体, 共获得505个F2隐性单株(即正常叶表型)。利用本实验室均匀分布于12条染色体上的617对SSR标记对亲本spl32和02428进行多态性分析, 其中共185个标记表现出明显的多态性, 多态率为30.08%。进一步利用在两亲本间表现出多态的引物, 扩增正常基因池和突变基因池, 发现第11染色体的标记RM21和RM206与突变体表型有明显的连锁。在定位区间周围, 进一步开发Indel分子标记13个, 其中4个有多态性(表3)。利用RM21、RM206以及4个Indel标记对505个正常绿叶的F2单株进行分析, 结果标记RM21、Ind-c、RM206和Ind-g分别有51、37、35及48个交换株, RM21交换株包含Ind-c交换株, Ind-g交换株包含RM206交换株, 且RM21、Ind-c交换株与RM206、Ind-g交换株互不重叠, 因此将Spl32(t)基因初步定位在Indel标记Ind-c与SSR标记 表2 Table 2 表2(Table 2)
Physical coordinates of primers were identified through BLAST searching in the Nipponbare rice genome (IRGSP-1.0). 物理位置是以引物序列在日本晴基因组(IRGSP-1.0)进行BLAST获得。
表3 定位基因Spl32(t)所用的4个Indel引物序列 Table 3 Four Indel primer sequences used for Spl32(t) mapping
Dangl JL, Dietrich RA, Richberg MH. Death don’t have no mercy: cell death programs in plant-microbe interactions. Plant Cell, 1966, 8: 1793-1807[本文引用:1][JCR: 9.251]
[5]
王建军, 朱旭东, 王林友, 张利华, 薛庆中, 何祖华. 水稻类病变突变体lrd40的抗病性与细胞学分析. 中国水稻科学, 2005, 19: 111-116Wang JJ, Zhu XD, Wang LY, Zhang LH, Xue QZ, He ZH. Disease resistance and cytological analyses on lesion resembling disease mutant lrd40 in Oryza sativa. Chin J Rice Sci, 2005, 19: 111-116 (in Chinese with Englinsh abstract)[本文引用:1][CJCR: 1.494]
[6]
陈析丰, 金杨, 马伯军. 水稻类病变突变体及抗病性的研究进展. 植物病理学报, 2011, 41: 1-9Chen XF, JinY, Ma BJ. Progress on the studies of rice lesion mimics and their resistant mechanism to the pathogens. Acta Phytopathol Sin, 2011, 41: 1-9 (in Chinese with Englinsh abstract)[本文引用:3][CJCR: 1.088]
[7]
Qiao YL, Jiang WZ, Lee JH, Park BS, Choi MS, Piao RH, Woo MO, Roh JH, Han LZ, Paek NC, Seo HS, Koh HJ. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit μ1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol, 2010, 185: 258-274[本文引用:5][JCR: 6.736]
[8]
Wu CJ, BordeosA, Madamba M R S, Baraoidan M, Ramos M, Wang G L, Leach J E, Leung H. Rice lesion mimic mutants with enhanced resistance to diseases. Mol Genet Genomics, 2008, 279: 605-619[本文引用:1][JCR: 2.881]
[9]
Zhong CY, JunC, Li RZ, Mei LG, HeiL, Gurdev SK, Wang GL. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight. Mol Plant-Microbe Interact, 2000, 13: 869-876[本文引用:1][JCR: 4.307]
[10]
Chen XF, HaoL, Pan JW, Zheng XX, Jiang GH, JinY, Gu ZM, QianQ, Zhai WX, Ma BJ. SPL5, a cell death and defense-related gene, encodes a putative splicing factor 3b subunit 3 (SF3b3) in rice. Mol Breed, 2012, 30: 939-949[本文引用:2][JCR: 3.251]
[11]
YamanouchiU, YanoM, Lin HX, AshikariM, YamadaK. A rice spotted leaf gene, Spl7, encodes a heat stress transcription factor protein. Proc Natl Acad Sci USA, 2002, 99: 7530-7535[本文引用:2][JCR: 9.737]
[12]
Zeng LR. Spotted leaf 11, a negative regulator of plant cell death and defense, encodes a U-Box/Armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell, 2004, 16: 2795-2808[本文引用:4][JCR: 9.251]
[13]
MoriM, TomitaC, SugimotoK, Hasegawa, HayashiN, Dubouzet J, Ochiai H, Sekimoto H, Hirochika H, Kikuchi S. Isolation and molecular characterization of a Spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol Biol, 2007, 63: 847-860[本文引用:3][JCR: 3.518]
[14]
WangL, PeiZ, TianY, HeC. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation. Mol Plant Microbe Interact, 2005, 18: 375-384[本文引用:2][JCR: 4.307]
[15]
ChernM, Fitzgerald HA, Canlas PE, Navarre DA, Ronald PC. Over expression of a rice NPR1 homolog leads to constitutive activation of defense response and hypersensitivity to light. Mol Plant-Microbe Interact, 2005, 18: 511-520[本文引用:1][JCR: 4.307]
Kim JA, ChoK, SinghR, Jung YH, Jeong SH, Kim SH, LeeJ, Cho YS, Agrawal GK, RakwalR, TamogamiS, KerstenB, Jeon JS, AnG, Jwa NS. Rice OsACDR1 (Oryza sativa accelerated cell death and resistance 1) is a potential positive regulator of fungal disease resistance. Mol Cells, 2009, 28: 431-439[本文引用:1][JCR: 2.21]
[18]
Jiang CJ, ShimonoM, MaedaS, InoueH, MoriM, HasegawaM, SuganoS, TakatsujiH. Suppression of the rice fatty-acid desaturase gene OsSSI2 enhances resistance to blast and leaf blight diseases in rice. Mol Plant Microbe Interact, 2009, 22: 820-829[本文引用:1][JCR: 4.307]
[19]
FujiwaraT, MaisonneuveS, IsshikiM, MizutaniM, ChenL, Wong HL, KawasakiT, ShimamotoK. Sekiguchi lesion gene encodes a cytochrome P450 monooxygenase that catalyzes conversion of tryptamine to serotonin in rice. J Biol Chem, 2010, 285: 11308-11313[本文引用:1][JCR: 4.651]
[20]
Sun CH, Liu LH, Tang JY, Lin AH, Zhang FT, FangJ, Zhang GF, Chu CC. RLIN1, encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice. Genet Genomics, 2011, 38: 29-37[本文引用:1][JCR: 2.397]
[21]
Jiao BB, Wang JJ, Zhu XD, Zeng LJ, LiQ, He ZH. A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice. Mol Plant, 2012, 5: 205-217[本文引用:3][JCR: 6.126][CJCR: 0.9487]
[22]
Tang JY, Zhu XD, Wang YQ, Liu LC, XuB, LiF, FangJ, Chu CC. Semi-dominant mutations in the CC-NB-LRR-type R gene, NLS1, lead to constitutive activation of defense responses in rice. Plant J, 2011, 66: 996-1007[本文引用:2][JCR: 6.582]
[23]
王丹. 水稻分蘖调控基因TE的功能分析和类病变突变体lms1的图位克隆. 中国农业科学院博士学位论文, 北京, 2012WangD. Functional Analysis of a Key Tillering Regulator TE and Map-based Cloning of Gene lms1 in Rice (Orzya sativa L. ). PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2012 (in Chinese with Englinsh abstract)[本文引用:2]
[24]
曹建. 水稻类病斑突变体的生理分析与LM基因的图位克隆. 中国农业科学院硕士学位论文, 北京, 2014CaoJ. Physiological Analysis of a Lesion Mimic Mutant and Map-based Cloning of Gene LM in Rice (Oryza sativa L. ). MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2014 (in Chinese with Englinsh abstract)[本文引用:1]
[25]
成晓越. 水稻类病变新基因CHL1 (chloroplastic-H2O2-induced Lesion 1) 的鉴定与克隆. 浙江师范大学硕士学位论文, 浙江杭州, 2013Cheng XY. Identification and Cloning of a Novel Rice Lesion Mimic Gene CHL1 (Chloroplastic-H2O2-induced Lesion 1). MS Thesis of Zhejiang Normal University, Zhejiang, China, 2013 (in Chinese with Englinsh abstract)[本文引用:1]
[26]
BuschgesR, HollricherK, PanstrugaR, SimonsG, WolterM, FrijtersA, Van DR, Vand er LT, DiergaardeP, GroenendijkJ, TopschS, VosP, SalaminiF, Schulze LP. The barley MLO gene: a novel control element of plant pathogen resistance. Cell, 1997, 88: 695-705[本文引用:1][JCR: 31.957]
[27]
Joergensen JH. Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica, 1992, 63: 141-152[本文引用:1][JCR: 1.643]
[28]
MizobuchiR, HirabayashiH, KajiR, NishizawaY, YoshimuraA, SatohH. Isolation and characterization of rice lesion-mimic mutants with enhanced resistance to rice blast and bacterial blight. Plant Sci, 2002, 163: 345-353[本文引用:2][JCR: 2.922]
[29]
Wu CJ, BordeosA, Madamba M R S, Baraoidan M, Ramos M, Wang G L, Leach J E, Leung H. Rice lesion mimic mutants with enhanced resistance to diseases. Mol Genet Genomics, 2008, 279: 605-619[本文引用:2][JCR: 2.881]
[30]
Huang QN, Shi YF, YangY, Feng BH, Wei YL, ChenJ, BaraoidanM, LeungH, Wu JL. Characterization and genetic analysis of a light-and temperature-sensitive spotted-leaf mutant in rice. J Integr Plant Biol, 2011, 53: 671-681[本文引用:2][JCR: 3.75][CJCR: 0.7555]
[31]
代高猛, 朱小燕, 李云峰, 凌英华, 赵芳明, 杨正林, 何光华. 水稻类病斑突变体spl31的遗传分析与基因定位. 作物学报, 2013, 39: 1223-1230Dai GM, Zhu XY, Li YF, Ling YH, Zhao FM, Yang ZL, He GH. Genetic analysis and fine mapping of a lesion mimic mutant spl31 in rice. Acta Agron Sin, 2013, 39: 1223-1230 (in Chinese with Englinsh abstract)[本文引用:1][CJCR: 1.667]
[32]
王建军, 朱旭东, 王友林, 张利华, 薛庆中, 何祖华. 水稻类病斑突变体的生理与遗传分析. 植物生理与分子生物学报, 2004, 30: 331-338Wang JJ, Zhu XD, Wang YL, Zhang LH, Xue QZ, He ZH. Physiological and genetic analysis of lesion mimic mutants in rice. J Plant Physiol Mol Biol, 2004, 30: 331-338 (in Chinese with Englinsh abstract)[本文引用:1][CJCR: 1.344]
[33]
YinZ, ChenJ, ZengL, GohM, LeungH, Khush GS, Wang GL. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight. Mol Plant Microbe Interact, 2000, 13: 869-876[本文引用:1][JCR: 4.307]
赵亚华. 生物化学实验技术教程. 广州: 华南理工大学出版社, 2000. pp 153-154Zhao YH. Biochemical Experimental Techniques Tutorial. Guangzhou: South China Science and Technology University Press, 2000. pp 153-154(in Chinese)[本文引用:1]
[36]
中国科学院上海植物生理研究所. 现代植物生理学实验指南. 北京: 科学出版社, 1999. pp 305-306Institute of PlantPhysiologyand Ecology, Shanghai Institutes of Biological Sciences, Chinese Academy ofSciences. Modern Laboratory Manual of Plant Physiology. Beijing: Science Press, 1999. pp 305-306(in Chinese)[本文引用:1]
[37]
杨敏文. 快速测定植物叶片叶绿素含量方法的探讨. 光谱实验室, 2002, 19: 478-481Yang MW. Study on rapid determination of chlorophyll content of leaves. Spectrographic Lab, 2002, 19: 478-481 (in Chinese with English abstract)[本文引用:1]
[38]
Lichtenthaler HK. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol, 1987, 148: 350-382[本文引用:1][JCR: 2.002]
[39]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1][JCR: 9.737]
[40]
Rogers SO, Bendich AJ. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol Biol, 1985, 5: 69-76[本文引用:1][JCR: 3.518]
[41]
Shen HC, Wang HM, Huang QN, XuX, Lu XG, Wu JL. Characterization and genetic analysis of a novel rice spotted-leaf mutant HM47 with broad-spectrum resistance to Xanthomonas oryzae pv. oryzae. Integr Plant Biol, 2013, 55: 473-483[本文引用:1][JCR: 3.75][CJCR: 0.7555]
[42]
梁颖, 李加纳, 唐章林, 谌利, 张学昆. 油菜光合生理指标与产量的关联分析. 西南农业大学学报, 1999, 21: 38-41LiangY, Li JN, Tang ZL, ChenL, Zhang XK. Correlative analysis of photosynthesis physiological targets and yield of rape. J Southwest Agric Univ, 1999, 21: 38-41 (in Chinese with English abstract)[本文引用:1][CJCR: 0.45]
[43]
王忠华. 植物类病变突变体的诱发与突变机制. 细胞生物学杂志, 2005, 27: 530-534Wang ZH. Induction and mutation mechanism of plant lesion mimic mutants. Chin J Cell Biol, 2005, 27: 530-534 (in Chinese with English abstract)[本文引用:1][CJCR: 0.3089]
[44]
Frye CA, TangD, Innes RW. Negative regulation of defense responses in plants by a conserved MAPKK kinase. Proc Natl Acad Sci USA, 2001, 98: 373-378[本文引用:1][JCR: 9.737]
[45]
金杨, 周丽芬, 陈析丰, 刘峰, 马伯军. 水稻类病变突变体spl5细胞坏死机制的分析. 浙江师范大学学报(自然科学版), 2009, 32: 329-330JinY, Zhou LF, Chen XF, LiuF, Ma BJ. Mechanisms of cell death in rice lesion mimic muant spl5. J Zhejiang Norm Univ (Nat Sci), 2009, 32: 326-331 (in Chinese with English abstract)[本文引用:1][CJCR: 0.3602]
[46]
虞玲锦. 一种水稻类病斑突变体生理分析与基因初定位, 南京林业大学硕士学位论文, 江苏南京, 2012Yu LJ. Physiological Analysis and Mapping of a Lesion Mimic Mutant in Rice (Oryza sativa L. ). MS Thesis of Nanjing Forestry University, Nanjing, China, 2012 (in Chinese with English abstract)[本文引用:1]
[47]
章琦. 水稻白叶枯病抗性基因鉴定进展及其利用. 中国水稻科学, 2005, 19: 453-459ZhangQ. Highlights in identification and application of resistance genes to bacterial blight. Chin J Rice Sci, 2005, 19: 453-459 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[48]
鄂志国, 张丽靖, 焦桂爱, 程本义, 王磊. 稻瘟病抗性基因的鉴定及利用进展. 中国水稻科学, 2008, 22: 533-540E ZG, Zhang L J, Jiao G A, Cheng B Y, Wang L. Highlights in identification and application of resistance genes to rice blast. Chin J Rice Sci, 2008, 22: 533-540 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[49]
林艳, 陈在杰, 田大刚, 杨广阔, 杨绍华, 刘华清, 陈松彪, 王锋. 水稻类病斑及早衰突变体lms1的鉴定及基因初步定位. 福建农业学报, 2014, 29(1): 29-34LinY, Chen ZJ, Tian DG, Yang GK, Yang SH, Liu HQ, Chen SB, WangF. Identification and gene mapping of a lesion mimic and senescence mutant lms1 in rice. Fujian J Agric Sci, 2014, 29(1): 29-34 (in Chinese with English abstract)[本文引用:1][CJCR: 0.7902]
 , 罗文龙
, 罗文龙







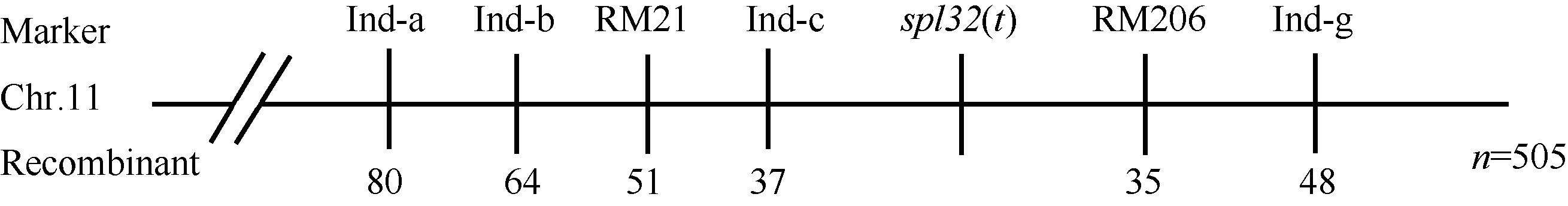

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}