关键词:小麦; 孢囊线虫; 白粉病; 抗病性 Development and Identification of Wheat Lines H3714 and H4058 Resistant to Cereal Cyst Nematode SUN Xi-Ying1,2, CUI Lei2,3, SUN Lei2, SUN Yan-Ling2, QIU Dan1,2, ZOU Jing-Wei1,2, WU Xiao-Fei2, WANG Xiao-Ming2, LI Hong-Jie2,* 1 College of Life Science and Technology, Hebei Normal University of Science and Technology, Qinhuangdao 066004, China
2 National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
3 Institute of Crop Science, Shanxi Academy of Agricultural Sciences, Taiyuan 030031, China
AbstractCereal cyst nematode (CCN, Heteroderaspp.) has occurred in major wheat ( Triticum aestivum L.) producing regions in China. Developing wheat cultivars resistant to CCN is limited due to the shortage of effective sources of resistance. Madsen, a winter wheat cultivar released in the Pacific Northwest Region of the USA, is highly resistant to CCN populations from China. However, it is difficult to use this cultivar in breeding programs because of its late heading and maturity. In the present study, Chinese winter wheat cultivars Yannong 21 and Jimai 19 were crossed and backcrossed to Madsen, and lines H3714 and H4058 were developed from BC1F4 progenies. Tests under both natural infestation and artificial inoculation conditions indicated that resistance of H3714 and H4058 to the H. avenae Xingyang population (pathotype Ha43) and the H. filipjevi Xuchang population (pathotype Hfc-1) from Henan province was superior to that of Yannong 21 and Jimai 19. Results of the field test inoculated with a mixture of Bgt isolates demonstrated that both lines showed adult plant resistance to powdery mildew, and line H4058 was also resistant to different Bgt isolates at the seedling stage. The heading date of these lines was comparable to that of Yannong 21 and Jimai 19, and obviously earlier than that of Madsen. The fragments of chromosome 2NS from Aegilops ventricosa were detected in lines H3714 and H4058 using a pair of chromosome-specific primers VENTRIUP-LN2 and the molecular markers Vlr2.6-3′-Vlr2.4-5′ and VRGA-F11-VRGA-R5 that were specific for Vrga1D gene from Ae. ventricosa. Results of Illumina iSelect 90K SNP assay showed that the two lines differed in their chromosome constitutions. Two thirds of the polymorphic SNPs were identical and the remaining ones were different among 4918 polymorphic SNPs between the two sib lines. The newly developed wheat lines H3714 and H4058 can be used as sources of resistance in developing CCN-resistant wheat cultivars.
Keyword: Triticum aestivum; Cereal cyst nematode; Powdery mildew; Resistance Show Figures Show Figures
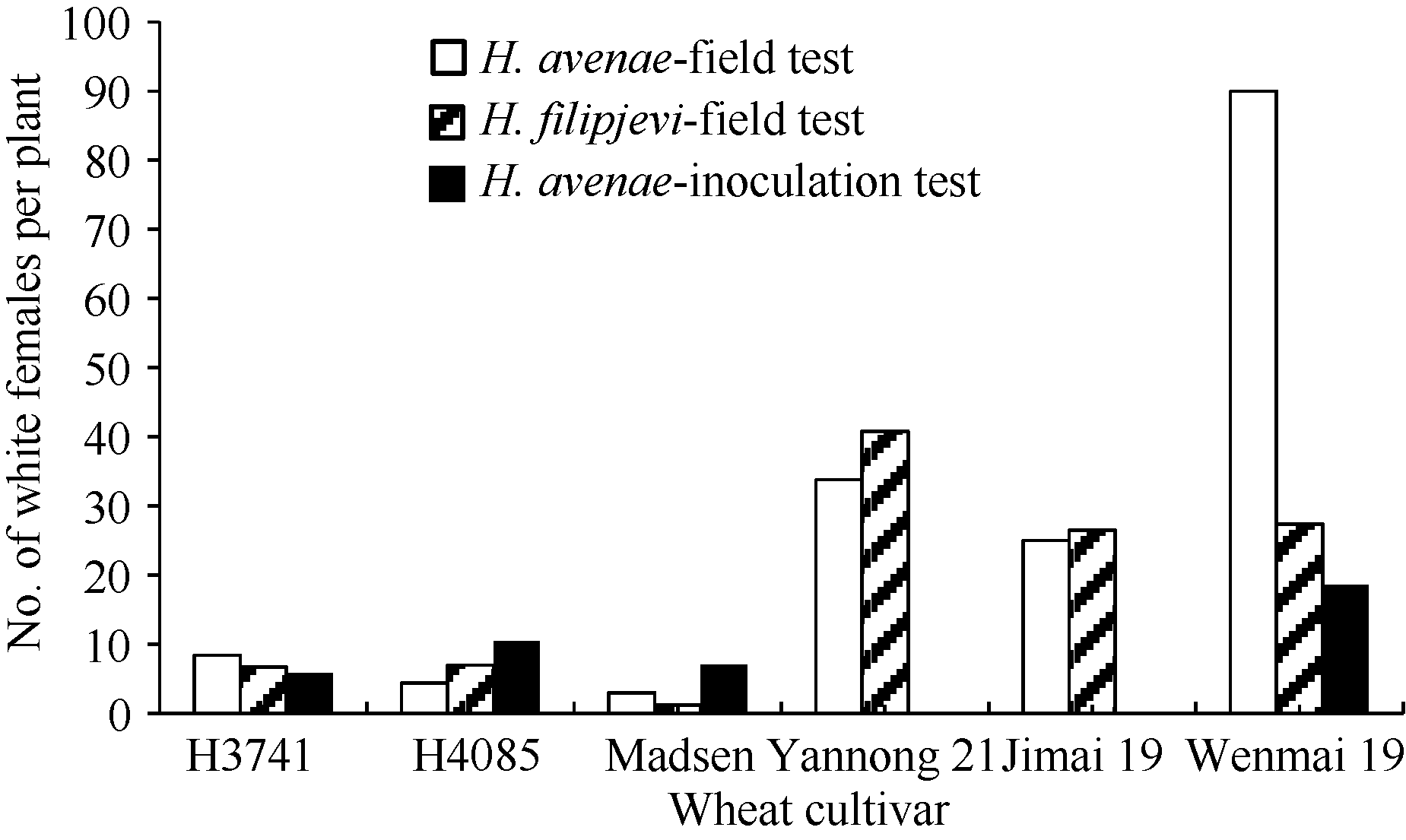
图1 田间病圃和接种条件下H3714和H4058及其亲本Madsen、烟农21和济麦19的单株孢囊数Fig. 1 Mean number of white females per plant developed on the roots of H3714, H4058 and their parental cultivars Madsen, Yannong 21, and Jimai 19 under natural infestation in the field tests and in the artificial inoculation test
表1 H3714、H4058和Madsen苗期和成株期对白粉病的反应 Table 1 Reactions of H3714, H4058, and Madsen to Blumeria graminis f. sp. tritici (Bgt) isolates at the seedling and adult plant stages
品系/品种 Line/cultivar
苗期接种Bgt菌株 Bgt isolates inoculated at the seedling stage
成株期 Adult plant stage
E09
E11
E20
E21
反应型 Infection type
最大病害严重度 Maximum disease severity
H3714
3
3
3
3
4
0
H4058
1
0
1
0
0
0
Madsen
3
3
4
3
7
85
中作9504 Zhongzuo 9504
4
4
4
4
8
100
表1 H3714、H4058和Madsen苗期和成株期对白粉病的反应 Table 1 Reactions of H3714, H4058, and Madsen to Blumeria graminis f. sp. tritici (Bgt) isolates at the seedling and adult plant stages
图2 利用偏凸山羊草2NS染色体特异引物VENTRIUP-LN2 (A)、Vlr2.6-3′ -Vlr2.4-5′ (B)和VRGA-F11-VRGA-R5 (C)对H3714和H4058及其亲本和对照材料的扩增M: 100 bp DNA ladder; 1: 偏凸山羊草; 2: VPM1; 3: Madsen; 4: H3714; 5: H4058; 6: 烟农21; 7: 济麦19; 8: 温麦19。 箭头示各标记扩增的特征带。Fig. 2 Amplification of H3714 and H4058 as well as their parental cultivars and the controls using primers Vlr2.6-3′ - Vlr2.4-5′ (A), VENTRIUP-LN2 (B), and VRGA-F11-VRGA-R5 (C) specific to chromosome 2NS from Aegilops ventricosaM: 100 bp DNA ladder; 1: Aegilops ventricosa; 2: VPM1; 3: Madsen; 4: H3714; 5: H4058; 6: Yannong 21; 7: Jimai 19; 8: Wenmai 19. Arrows indicate the diagnostic bands amplified by each pair of primers.
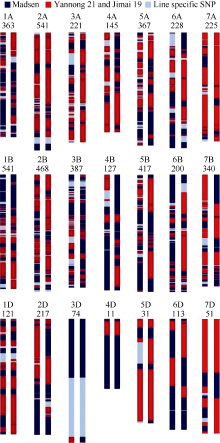
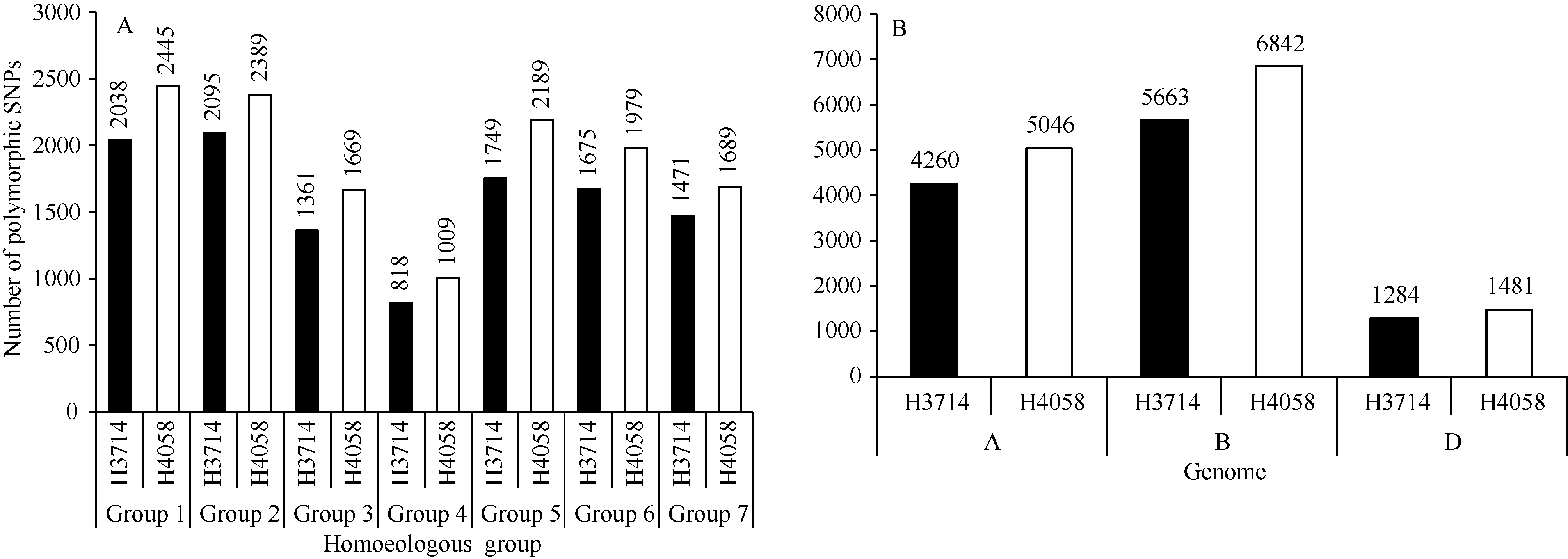
图3 多态性SNP位点在H3714和H4058部分同源群(A)和基因组(B)上的分布Fig. 3 Distributions of polymorphic SNPs on different homoeologous groups (A) and genomes (B) in lines H3714 and H4058
图4 根据多态性SNP绘制的H3714和H4058的染色体构成图相同编号并列的2条染色体, 左侧为H3714, 右侧为H4058; 染色体编号下的数字为SNP数。Fig. 4 A diagram of chromosomal compositions for H3714 and H4058 based on the polymorphic SNPsThe two chromosomes under the same codes belong to H3714 (left) and H4058 (right). The number of SNPs used to construct the genetic maps of individual chromosomes is indicated under the chromosome codes.
Peng DL, Nicol JM, Li HM, Hou SY, Li HX, Chen SL, MaP, Li HL, Riley IT. Current knowledge of cereal cyst nematode (Heterodera avenae) on wheat in China. In: Riley I T, Nicol J M, Dababat A A, eds. Cereal Cyst Nematodes: Status, Research and Outlook. Ankara, Turkey: CIMMYT Press, 2009. pp 29-34[本文引用:1]
[2]
李惠霞, 柳永娥, 魏庄, 李敏全. 西藏和新疆自治区麦田禾谷孢囊线虫的检测. 见: 廖金铃, 彭德良, 段玉玺, 简恒, 李红梅(主编). 中国线虫学研究, 第四卷, 北京: 中国农业科学技术出版社, 2012. pp 164-165Li HX, Liu YE, WeiZ, Li MQ. The detection of Heterodera avenae from the cereal field in autonomous region of Tibet and Xinjiang. In: Liao J L, Peng D L, Duan Y X, Jian H, Li H M, eds. Nematology Research in China, Vol. 4. Beijing: China Agricultural Science and Technology Press, 2012. pp 164-165(in Chinese)[本文引用:1]
[3]
刘刚. 农业部种植业管理司提醒跨区收割可传带小麦孢囊线虫. 北京农业, 2009, (9): 42LiuG. The Ministry of Agriculture Planting Industry Management department reminds that crop harvesting across regions can spread cereal cyst nematode. Beijing Agric, 2009, (9): 42[本文引用:1][CJCR: 0.1475]
[4]
Li HJ, CuiL, Li HL, Wang XM, Murray TD, Conner RL, Wang LW, GaoX, SunY, Sun SC, Tang WH. Effective resources in wheat and Wheat-Thinopyrum derivatives for resistance to Heterodera filipjevi in China. Crop Sci, 2012, 52: 1209-1212[本文引用:3][JCR: 1.513]
[5]
赵洪海, 杨远永, 彭德良. 山东省主要小麦品种对禾谷孢囊线虫抗性的初步评价. 山东农业科学, 2012, 44(2): 80-83Zhao HH, Yang YY, Peng DL. Preliminary evaluation on resistance of main wheat cultivars from Shand ong province to cereal cyst nematode. Shand ong Agric Sci, 2012, 44(2): 80-83 (in Chinese)[本文引用:1][CJCR: 0.58]
[6]
李秀花, 马娟, 高波, 王容燕, 陈书龙. 部分国内外小麦种质资源对燕麦孢囊线虫的抗病性. 麦类作物学报, 2013, 33: 1277-1283Li XH, MaJ, GaoB, Wang RY, Chen SL. Resistance of wheat cultivars or germplasm lines to Heterodera avenae. J Triticeae Crops, 2013, 33: 1277-1283 (in Chinese with English abstract)[本文引用:1][CJCR: 1.007]
[7]
邢小萍, 袁虹霞, 孙君伟, 张洁, 孙炳剑, 李洪连. 河南省小麦主推品种对2种禾谷孢囊线虫的抗性及其评价方法. 作物学报, 2014, 40: 805-815Xing XP, Yuan HX, Sun JW, ZhangJ, Sun BJ, Li HL. Resistance to two species of cereal cyst nematode and evaluation methods in major wheat cultivars from Henan province, China. Acta Agron Sin, 2014, 40: 805-815 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[8]
刘炳良, 孙成刚, 王暄, 向桂林, 宋志强, 高菲菲, 李红梅. 小麦品种对禾谷孢囊线虫(Heterodera avenae)江苏沛县群体的抗性鉴定. 麦类作物学报, 2012, 32: 563-568Liu BL, Sun CG, WangX, Xiang GL, Song ZQ, Gao FF, Li HM. Evaluation of the resistance of wheat cultivars to Peixian population of Heterodera avenae from Jiangsu province, China. J Triticeae Crops, 2012, 32: 563-568 (in Chinese with English abstract)[本文引用:1][CJCR: 1.007]
[9]
代君丽, 崔磊, 刘珂, 宗莹莹, 袁虹霞, 邢小萍, 李洪杰, 李洪连. 小麦品种太空6号对Heterodera avenae郑州群体的抗性遗传分析. 作物学报, 2013, 39: 642-648Dai JL, CuiL, LiuK, Zong YY, Yuan HX, Xing XP, Li HJ, Li HL. Genetic analysis of common wheat cultivar Taikong 6 for resistance to Heterodera avenae Zhengzhou population. Acta Agron Sin, 2013, 39: 642-648 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[10]
宗莹莹, 代君丽, 袁虹霞, 邢小萍, 孙炳剑, 李洪连. 普通小麦品种中育6号对两种禾谷孢囊线虫的抗性遗传分析. 麦类作物学报, 2013, 33: 249-254Zong YY, Dai JL, Yuan HX, Xing XP, Sun BJ, Li HL. Genetic analysis of resistance to cereal cyst nematode in common wheat variety Zhongyu 6. J Triticeae Crops, 2013, 33: 249-254 (in Chinese with English abstract)[本文引用:1][CJCR: 1.007]
[11]
袁虹霞, 张福霞, 张佳佳, 侯兴松, 李洪杰, 李洪连. CIMMYT小麦种质资源对菲利普孢囊线虫(Heterodera filipjevi)河南许昌群体的抗性. 作物学报, 2011, 37: 1956-1966Yuan HX, Zhang FX, Zhang JJ, Hou XS, Li HJ, Li HL. Resistance of CIMMYT wheat germplasm to Heterodera filipjevi Xuchang population from Henan province, China. Acta Agron Sin, 2011, 37: 1956-1966 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[12]
高秀, 崔磊, 李洪连, 王晓鸣, 唐文华, Conner RL, 林小虎, 李洪杰. 硬粒小麦品种Waskana和Waskowa对禾谷孢囊线虫(Heterodera filipjevi和H. avenae)的抗性. 作物学报, 2012, 38: 571-577GaoX, CuiL, Li HL, Wang XM, Tang WH, Conner RL, Lin XH, Li HJ. Resistance of Triticum durum cultivars Waskana and Waskowa to cereal cyst nematode, Heterodera filipjevi and H. avenae. Acta Agron Sin, 2012, 38: 571-577 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[13]
武小菲, 李洪杰, 王晓鸣, 陈怀谷, 徐世昌, 刘太国. 28份人工合成小麦对禾谷孢囊线虫、纹枯病、条锈病和叶锈病的抗性. 植物遗传资源学报, 2013, 14: 1221-1226Wu XF, Li HJ, Wang XM, Chen HG, Xu SC, Liu TG. Reactions of synthetic wheat to cereal cyst nematode, sharp eyespot, stripe rust, and leaf rust. J Plant Genet Resour, 2013, 14: 1221-1226 (in Chinese with English abstract)[本文引用:1][CJCR: 1.1628]
[14]
张佳佳, 袁虹霞, 张瑞奇, 邢小萍, 代君丽, 牛吉山, 李洪连, 陈佩度. 普通小麦-簇毛麦种质对菲利普孢囊线虫的抗性分析. 作物学报, 2012, 38: 1969-1976Zhang JJ, Yuan HX, Zhang RQ, Xing XP, Dai JL, Niu JS, Li HL, Chen PD. Analysis of resistance to Heterodera filipjevi in Triticum aestivum-Dasypyrum villosum germplasm. Acta Agron Sin, 2012, 38: 1969-1976 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[15]
Allan RE, Peterson Jr C J, Rubenthaler G L, Line R F, Roberts D E. Registration of Madsen wheat (Reg. No. 746). Crop Sci, 1989, 29: 1575-1576[本文引用:4][JCR: 1.513]
[16]
HelgueraM, Khan IA, KolmerJ, LijavetzkyD, Zhong-qi L, Dubcovsky J. PCR assays for the Lr37-Yr17-Sr38 cluster of rust resistance genes and their use to develop isogenic hard red spring wheat lines. Crop Sci, 2003, 43: 1839-1847[本文引用:6][JCR: 1.513]
[17]
WangS, WongD, ForrestK, AllenA, ChaoS, Huang BE, MaccaferriM, SalviS, Milner SG, CattivelliL, Mastrangelo AM, WhanA, StephenS, BarkerG, WiesekeR, PlieskeJ, International Wheat Genome Sequencing Consortium, Lillemo M, Mather D, Appels R, Dolferus R, Brown-Guedira G, Korol A, Akhunova A R, Feuillet C, Salse J, Morgante M, Pozniak C, Luo M C, Dvorak J, Morell M, Dubcovsky J, Ganal M, Tuberosa R, Lawley C, Mikoulitch I, Cavanagh C, Edwards K J, Hayden M, Akhunov E. Characterization of polyploid wheat genomic diversity using a high-density 90, 000 single nucleotide polymorphism array. Plant Biotechnol J, 2014, 12: 787-796[本文引用:4][JCR: 6.279]
[18]
MaiaN. Obtention de bles tenders resistants au pietin-verse par croisements interspecifiques bles × Aegilops. CR Acad Agric (Fr. ), 1967, 53: 149-154 (in French with English abstract)[本文引用:1]
[19]
Yuan HX, Sun JW, Yang WX, Xing XP, Wang ZY, Riley IT, Li HL. New pathotypes of Heterodera avenae (cereal cyst nematode) from winter wheat in Zhengzhou, Henan, China. Australas Plant Pathol, 2010, 39: 107-111[本文引用:1]
[20]
Li HL, Yuan HX, Sun JW, FuB, Nian GL, Hou XS, Xing XP, Sun BJ. First record of the cereal cyst nematode Heterodera filipjevi in China. Plant Dis, 2010, 94: 1505[本文引用:1][JCR: 2.455]
[21]
Nicol JM, OgbonnayaF, Singh AK, Bishnoi SP, Kanwar RS, Li HL, Chen SL, Peng DL, BolatN, ŞahinE, ElekcioğluİH. Current global knowledge of the usability of cereal cyst nematode resistant bread wheat germplasm through international germplasm exchange and evaluation. In: Riley I T, Nicol J M, Dababat A A, eds. Cereal Cyst Nematodes: Status, Research and Outlook. Ankara, Turkey: CIMMYT Press, 2009. pp 149-153[本文引用:2]
[22]
李洪杰, 王晓鸣, 宋凤景, 伍翠平, 武小菲, 张宁, 周阳, 张学勇. 中国小麦品种对白粉病的抗性反应与抗病基因检测. 作物学报, 2011, 37: 943-954Li HJ, Wang XM, Song FJ, Wu CP, Wu XF, ZhangN, ZhouY, Zhang XY. Response to powdery mildew and detection of resistance genes in wheat cultivars from China. Acta Agron Sin, 2011, 37: 943-954 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[23]
SeahS, SpielmeyerW, JahierJ, SivasithamparamK, Lagudah ES. Resistance gene analogs within an introgressed chromosomal segment derived from Triticum ventricosum that confers resistance to nematode and rust pathogens in wheat. Mol Plant- Microbe Interact, 2000, 13: 334-341[本文引用:1][JCR: 4.307]
[24]
SeahS, BarianaH, JahierJ, SivasithamparamK, Lagudah ES. The introgressed segment carrying rust resistance genes Yr17, Lr37 and Sr38 in wheat can be assayed by a cloned disease resistance gene-like sequence. Theor Appl Genet, 2001, 102: 600-605[本文引用:3][JCR: 3.658]
[25]
Fang TL, Campbell KG, Liu ZY, Chen XM, Wan AM, LiS, Liu ZJ, Cao SH, Chen YH, Bowden RL, Carver BF, Yan LL. Stripe rust resistance in the wheat cultivar Jagger is due to Yr17 and a novel resistance gene. Crop Sci, 2011, 51: 2455-2465[本文引用:2][JCR: 1.513]
[26]
van BerlooR. GGT 2. 0: Versatile software for visualization and analysis of genetic data. J Hered, 2008, 99: 232-236[本文引用:1][JCR: 1.995]
[27]
BekalS, JahierJ, RivoalR. Host responses of Triticeae to species of the cereal cyst nematode complex in relation to breeding resistant durum wheat. Fundam Appl Nematol, 1998, 21: 359-370[本文引用:1]
[28]
Bariana HS, McIntosh R A. Characterisation and origin of rust and powdery mildew resistance genes in VPM1 wheat. Euphytica, 1994, 76: 53-61[本文引用:2][JCR: 1.643]
[29]
JahierJ, AbelardP, TanguyM, DedryverF, RivoalR, KhatkarS, Bariana HS, KoebnerR. The Aegilops ventricosa segment on chromosome 2AS of the wheat cultivar ‘VPM1’ carries the cereal cyst nematode resistance gene Cre5. Plant Breed, 2001, 120: 125-128[本文引用:1][JCR: 1.175]
[30]
McIntosh RA, Wellings CR, Park RF. Wheat Rusts, an Atlas of Resistance Genes. CSIRO, Melbourne, Australia, 1995. [本文引用:1]
[31]
TakeuchiT, MunekataS, SuzukiT, SendaK, HoritaH, ArakiK, AsayamaS, SatoM. Breeding wheat lines resistant to wheat yellow mosaic virus and localization of the resistance gene (YmMD) derived from wheat cultivar 'Madsen'. Breed Res, 2010, 12: 1-8[本文引用:1]
[32]
Williamson VM, ThomasV, FerrisH, DubcovskyJ. An Aegilops ventricosa translocation confers resistance against root-knot nematodes to common wheat. Crop Sci, 2013, 53: 1412-1418[本文引用:1][JCR: 1.513]
[33]
Dyck PL, Lukow OM. The genetic analysis of two interspecific sources of leaf rust resistance and their effect on the quality of common wheat. Can J Plant Sci, 1988, 68: 633-639[本文引用:1][JCR: 0.716]
[34]
CaneK, Sharp PJ, Eagles HA, Eastwood RF, Hollamby GJ, KuchelH, Lu MQ, Martin PJ. The effects on grain quality traits of a grain serpin protein and the VPM1 segment in southern Australian wheat breeding. Aust J Agric Res, 2008, 59: 883-890[本文引用:1][JCR: 1.328]
[35]
RobertO, AbelardC, DedryverF. Identification of molecular markers for the detection of the yellow rust resistance gene Yr17 in wheat. Mol Breed, 1999, 5: 167-175[本文引用:1][JCR: 3.251]
[36]
AmbrozkováM, DedryverF, DumalasováV, HanzalováA, BartošP. Determination of the cluster of wheat rust resistance genes Yr17, Lr37 and Sr38 by a molecular marker. Plant Prot Sci, 2002, 38: 41-45[本文引用:1]
[37]
Hanzalová, A, Sumíková T, Bartoš P. Determination of leaf rust resistance genes Lr10Lr26 and Lr37 by molecular markers in wheat cultivars registered in the Czech Republic. Czech J Genet Plant Breed, 2009, 45: 79-84[本文引用:1][JCR: 0.385]
[38]
李峰奇, 韩德俊, 魏国荣, 曾庆东, 康振生. 黄淮麦区小麦品种Lr37-Yr17-Sr38基因簇的分子检测. 西北农林科技大学学报(自然科学版), 2009, 37(3): 151-158Li FQ, Han DJ, Wei GR, Zeng QD, Kang ZS. Identification of Lr37-Yr17-Sr38 in wheat cultivars of Huanghuai wheat region using molecular markers. J Northwest A&F Univ (Nat Sci Edn), 2009, 37(3): 151-158 (in Chinese with English abstract)[本文引用:1]
[39]
RobertO, DedryverF, LeconteM, Rolland B, de Vallavieille-Pope C. Combination of resistance tests and molecular tests and molecular tests to postulate the yellow rust resistance gene Yr17 in bread wheat lines. Plant Breed, 2000, 119: 467-472[本文引用:1][JCR: 1.175]
[40]
WurschumT, Langer SM, Longin C F H, Korzun V, Akhunov E, Ebmeyer E, Schachschneider R, Schacht J, Kazman E, Reif J C. Population structure, genetic diversity and linkage disequilibrium in elite winter wheat assessed with SNP and SSR markers. Theor Appl Genet, 2013, 126: 1477-1486[本文引用:1][JCR: 3.658]
[41]
Cavanagh CR, ChaoS, WangS, Huang BE, StephenS, KianiS, ForrestK, SaintenacC, Brown-Guedira G L, Akhunova A, See D, Bai G, Pumphrey M, Tomar L, Wong D, Kong S, Reynolds M, da Silva M L, Bockelman H, Talbert L, Anderson J A, Dreisigacker S, Baenziger S, Carter A, Korzun V, Morrell P L, Dubcovsky J, Morell M K, Sorrells M E, Hayden M J, Akhunov E. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat land races and cultivars. Proc Natl Acad Sci USA, 2013, 110: 8057-8062[本文引用:1][JCR: 9.737]
 , 崔磊
, 崔磊





{kind=link}
{kind=link}
{kind=link}
{kind=link}