关键词:抽穗期; 染色体片段代换系; 多穗; 多粒; 高叶绿素含量 Identification and Morphological Analysis of a Rice Chromosome Segment Substitution Line Carrying a Major Effect Gene for Late Heading Date and Mapping of Ehd4-2 XIANG Jia, LI Yan, FAN Ya-Wei, XU Jun-Hong, ZHENG Li-Yuan, HE Guang-Hua, YANG Zheng-Lin, WANG Nan, ZHAO Fang-Ming* Rice Research Institute, Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Chongqing 400716, China
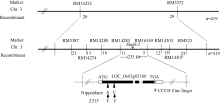
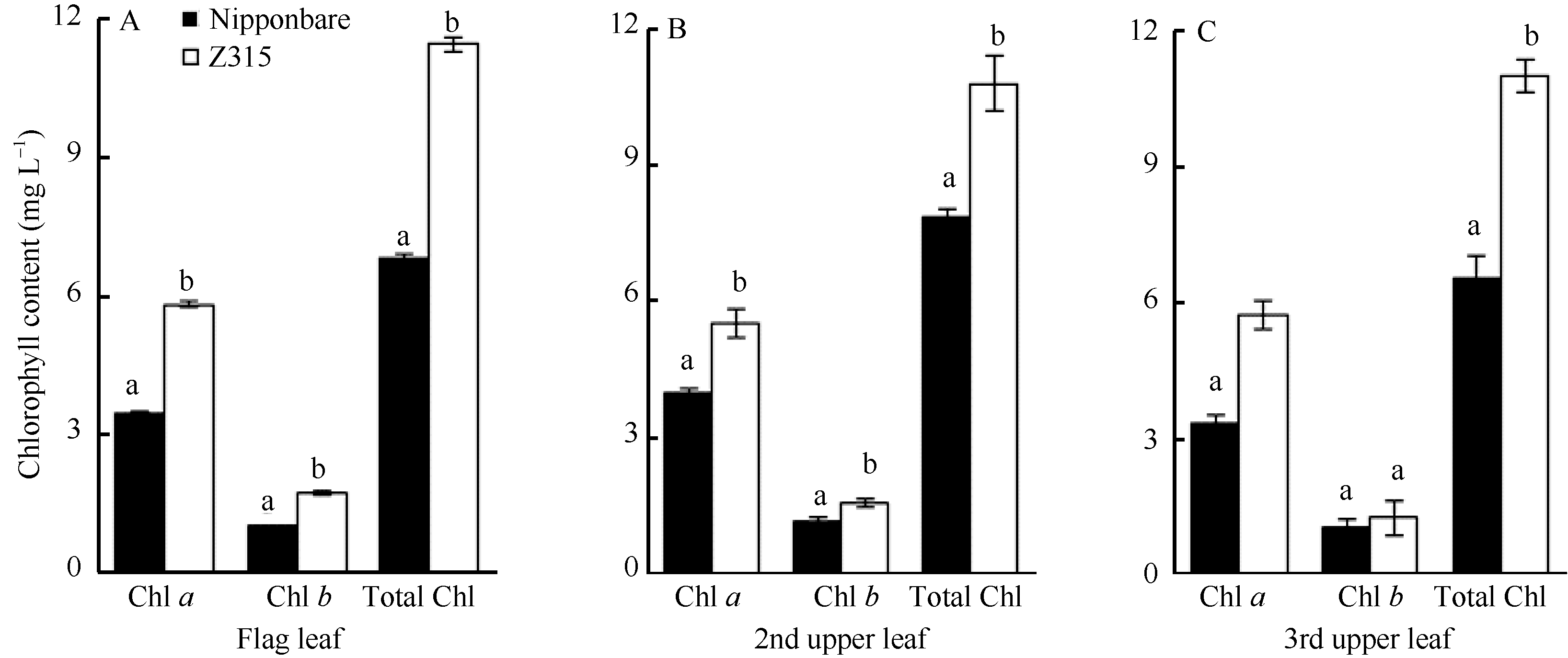
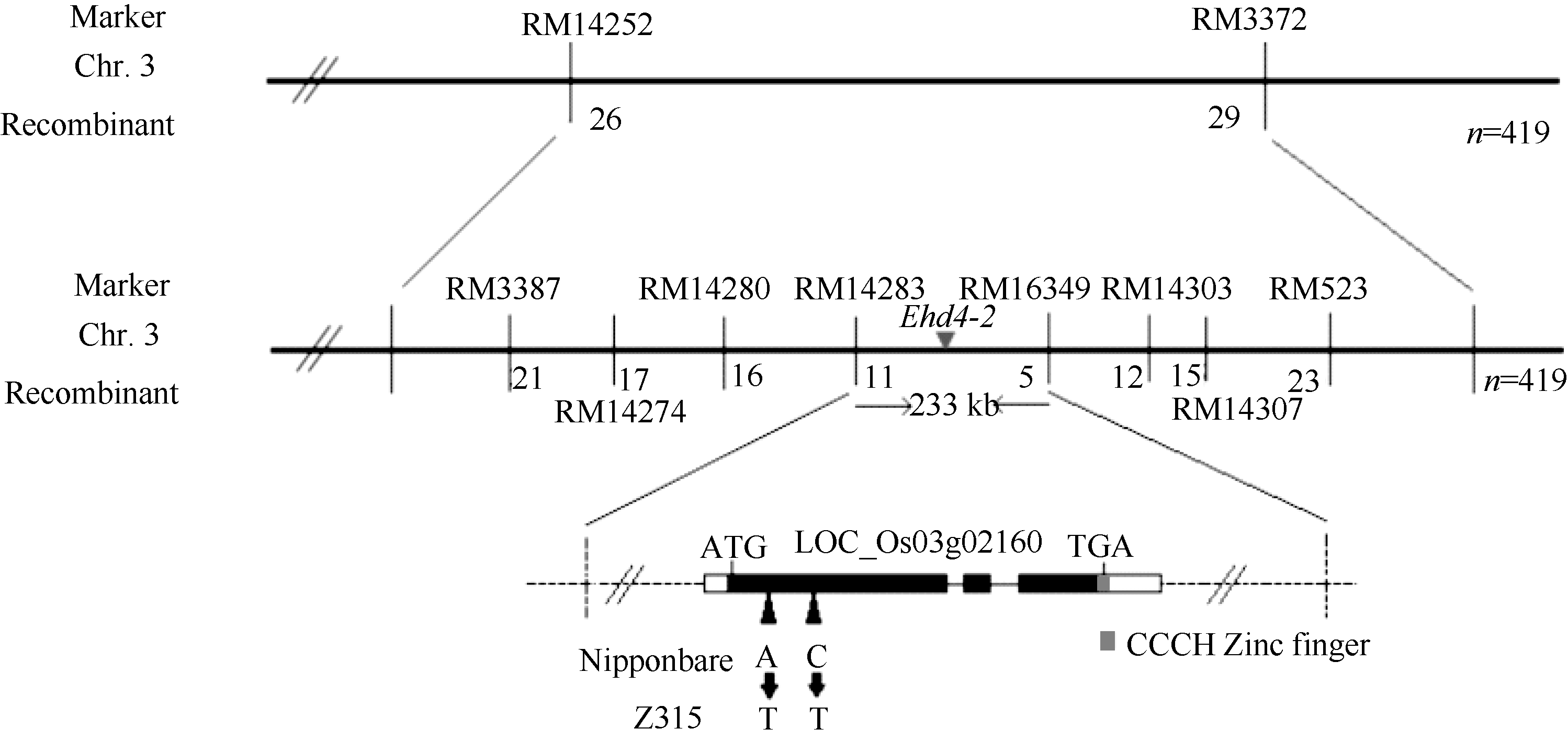
AbstractHeading date is an important agronomic trait deciding planting area and seasonal adaptability of rice varieties. Identification of genes for heading date is important for rice production. In this study, a novel chromosome segment substitution line (CSSL) named Z315, carrying a single gene for late heading date, was identified by advanced backcrosses between the recipient Nipponbare and donor Xihui 18 and SSR marker-assisted selection. Z315 carried five substitution segments from Xihui 18. They were distributed on chromosomes 1, 3, 6, and 7. The average length of these substitution segments is 7.39 Mb. The chlorophyll content, plant height, panicle length, length of the 1st internode, length of the 2nd and 3rd upper leaf, number of efficient panicles, number of spikilets per panicle and number of grains per panicle in Z315 were significantly higher than those in Nipponbare, suggesting that QTLs (quantitative trait locus) for these traits might be carried on the chromosome substitution segments. Furthermore, F1 and F2 populations from cross of Nipponbare and Z315 were used for genetic analysis and gene mapping for the late heading date. Genetic analysis showed that the late heading date phenotype was controlled by a single recessive nuclear gene, which was mapped between SSR markers RM14283 and RM6349 on chromosome 3, with a physical distance of 233 kb. By gene prediction and candidate gene sequencing, a gene (LOC_Os03g02160) related to heading date was found existing differences between Nipponbare and Z315. The gene encoded zinc finger protein, which might be an allele of Ehd4, named Ehd4-2. The results in the paper have important applications for further dissection of other QTLs in Z315 and breeding for gene pyramiding because chromosome segment substitution lines have the same genetic background besides substitution segments compared with the recipient parent.
Keyword:Heading date; Chromosome segment substitution lines; More panicles; More grains; High content of chlorophyll Show Figures Show Figures
图2 日本晴和Z315的表型. A: 抽穗期的日本晴和Z315; B: 成熟期的日本晴和Z315。Fig. 2 Phenotypes of Nipponbare and Z315. A: phenotypes of Nipponbare and Z315 at heading stage; B: phenotypes of Nipponbare and Z315 at maturity stage.
表1 Table 1 表1(Table 1)
表1 日本晴与Z315的株高、穗长和节间长 Table 1 Plant height, length of panicle, and internode of Nipponbare and Z315
材料 Material
株高 Plant height (cm)
穗长 Panicle length (cm)
倒一节间长 1st internode length (cm)
倒二节间长 2nd internode length (cm)
倒三节间长 3rd internode length (cm)
日本晴Nipponbare
104.89± 5.93 a
18.37± 1.00 a
19.55± 3.10 a
16.00± 3.64 a
12.37± 2.32 a
Z315
121.18± 5.71 b
19.47± 1.02 b
23.72± 3.70 b
16.71± 2.37 a
14.03± 2.71 a
Values followed by a and b are significantly different at P< 0.05 between two materials. 标以a和b的值在两材料间差异显著(P< 0.05).
表1 日本晴与Z315的株高、穗长和节间长 Table 1 Plant height, length of panicle, and internode of Nipponbare and Z315
表2 Table 2 表2(Table 2)
表2 成熟期日本晴与Z315的叶片形态 Table 2 Leaf morphology of Nipponbare and Z315 at maturity stage
材料 Material
剑叶长 Length of flag leaf (cm)
剑叶宽 Width of flag leaf (cm)
倒二叶长 Length of 2nd upper leaf (cm)
倒二叶宽 Width of 2nd upper leaf (cm)
倒三叶长 Length of 3rd upper leaf (cm)
倒三叶宽 Width of 3rd upper leaf (cm)
日本晴Nipponbare
25.81± 7.58 a
1.08± 0.22 a
30.90± 4.84 a
1.02± 0.29 a
36.13± 6.74 a
0.92± 0.25 a
Z315
29.76± 6.07 a
1.10± 0.26 a
33.92± 4.78 b
0.94± 0.19 a
41.24± 5.18 b
0.86± 0.18 a
Values followed by a and b are significantly different at P< 0.05 between two materials. 标以a和b的值在两材料间差异显著(P< 0.05).
表2 成熟期日本晴与Z315的叶片形态 Table 2 Leaf morphology of Nipponbare and Z315 at maturity stage
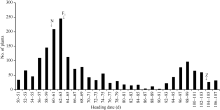
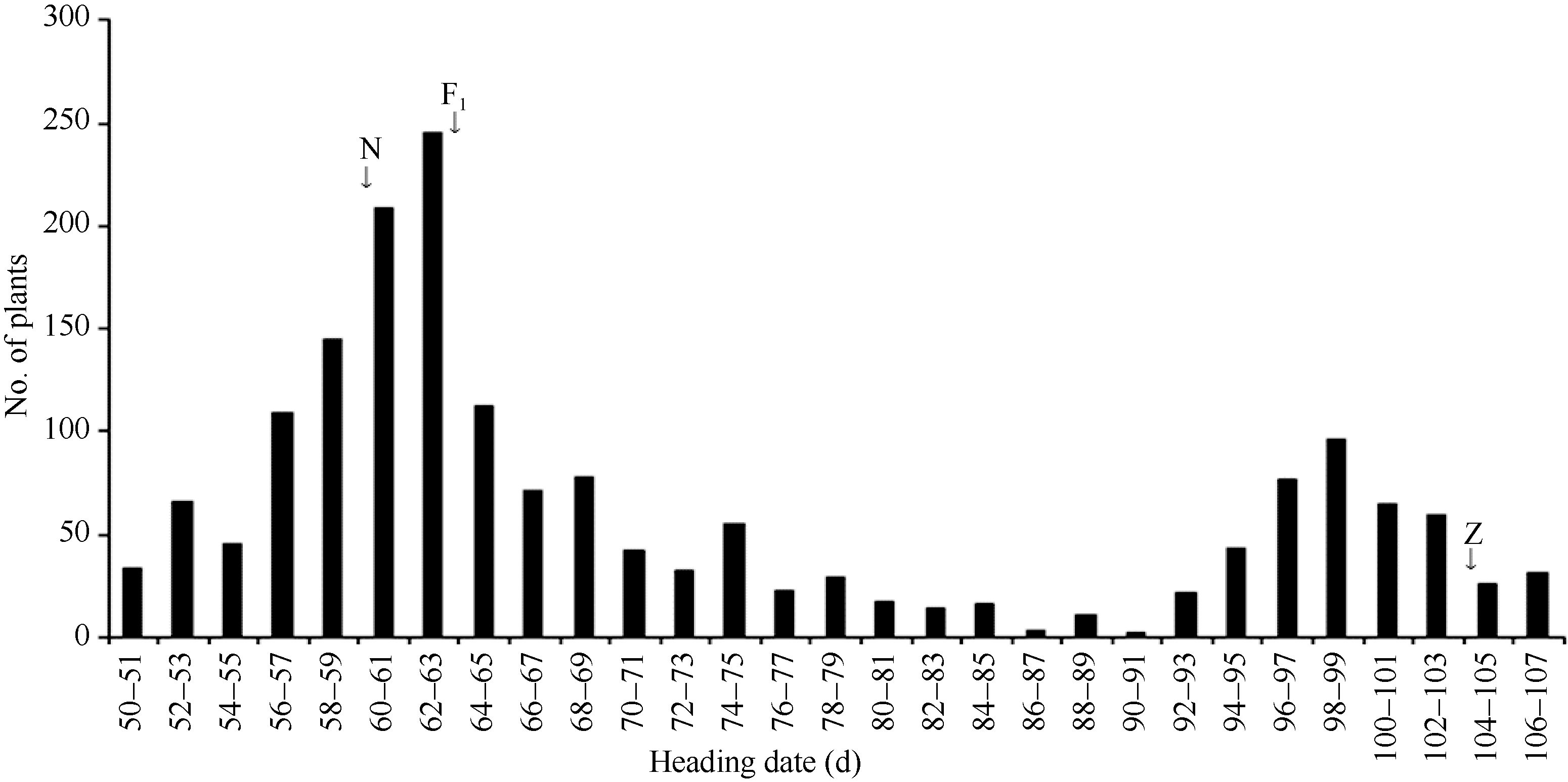
图4 日本晴和Z315杂交的F2群体抽穗天数分布. N: 日本晴; Z: 代换系Z315; F1: 日本晴与Z315杂交的F1群体。Fig. 4 Distribution of heading date of F2 population from cross of Nipponbare and Z315. N: recipient Nipponbare; Z: CSSL Z315; F1: F1 population from cross of Nipponbare/Z315.
Khush GS. Productivity improvements in rice. Nutr Rev, 2003, 61: 8114-8116[本文引用:1][JCR: 4.597]
[2]
Khush GS. Challenges for meeting the global food and nutrient needs in the new millennium. Proc Nutr Soc, 2001, 60: 15-26[本文引用:1]
[3]
Du MM, WangC. Progress on mapping of quantitative trait loci in crops. J Northeast Agric Univ, 2008, 15: 63-67[本文引用:1]
[4]
Liao CY, WuP, HuB, Yi KK. Effects of genetic background and environment on QTLs and epistasis for rice (Oryza sativa L. ) panicle number. Theor Appl Genet, 2001, 103: 104-111[本文引用:1][JCR: 3.658]
[5]
魏祥进, 徐俊峰, 江玲, 王洪俊, 周振玲, 翟虎渠, 万建民. 我国水稻主栽品种抽穗期多样性的遗传分析. 作物学报, 2012, 38: 10-22Wei XJ, Xu JF, JiangL, Wang HJ, Zhou ZL, Zhai HQ, Wan JM. Genetic analysis for the diversity of heading date of cultivated rice in China. Acta Agron Sin, 2012, 38: 10-22 (in Chinese with English abstract)[本文引用:1]
[6]
YanoM, KatayoseY, AshikariM, YamanouchiU, MonnaL, FuseT, BabaT, YamamotoK, UmeharaY, NagamuraY, SasakiT. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell, 2000, 12: 2473-2484[本文引用:2][JCR: 9.251]
[7]
YamamotoT, Lin HX, SasakiT, YanoM. Identification of heading date quantitative trait locus Hd6 and characterization of its epistatic interactions with Hd2 in rice using advanced backcross progeny. Genetics, 2000, 154: 885-891[本文引用:2][JCR: 4.389]
[8]
TakahashiY, ShomuraA, SasakiT, YanoM. Hd6, a rice quantitative trait locus involved in photoperiod sensitivity, encodes the a subunit of protein kinase CK2. Proc Natl Acad Sci USA, 2001, 98: 7922-7927[本文引用:3][JCR: 9.737]
[9]
KojimaS, TakahashiY, KobayashiY, MonnaL, SasakiT, ArakiT, YanoM. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day condition. Plant Cell Physiol, 2002, 43: 1096-1105[本文引用:4][JCR: 4.134]
[10]
DoiK, IzawaT, FuseT, YamanouchiU, KuboT, ShimataniZ, YanoM, YoshimuraA. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes & Development, 2004, 18: 926-936[本文引用:2]
[11]
MatsubaraK, YamanouchiU, Wang ZX, MinobeY, IzawaT, YanoM. Ehd2, a rice ortholog of the maize INDETERMINATE1 gene, promotes flowering by up-regulating Ehd1. Plant Physiol, 2008, 148: 1425-1435[本文引用:3][JCR: 6.555]
[12]
MatsubaraK, YamanouchiU, NonoueY, SugimotoK, Wang ZX, MinobeY, YanoM. Ehd3, encoding a plant homeodomain finger-containing protein, is a critical promoter of rice flowering. Plant J, 2011, 66: 603-612[本文引用:3][JCR: 6.582]
[13]
GaoH, Zheng XM, Fei GL, ChenJ, Jin MN, Ren YL, Wu WX, Zhou KN, Sheng PK, ZhouF, JiangL, WangJ, ZhangX, Guo XP, Wang JL, Cheng ZJ, Wu CY, Wang HY, Wan JM. Ehd4 encodes a novel and Oryza-genus-specific regulator of photoperiodic flowering in rice. PLoS Genet, 2013, 9: e1003281[本文引用:6][JCR: 8.517]
[14]
Wu WX, Zheng XM, Lu GW, Zhong ZZ, GaoH, Chen LP, Wu CY, Wang HJ, WangQ, Zhou KN, Wang JL, Wu FQ, ZhangX, Gu XP, Cheng ZJ, Lei CL, Lin QB, JiangL, Wang HY, GeS, Wan JM. Association of functional nucleotide polymorphisms at DTH2 with the northward expansion of rice cultivation in Asia. Proc Natl Acad Sci USA, 2013, 110: 2775-2780[本文引用:3][JCR: 9.737]
[15]
LiJ, Chu HW, Zhang YH, Mou TM, Wu CY, Zhang QF, XuJ. The rice HGW gene encodes a ubiquitin-associated (UBA) domain protein that regulates heading date and grain weight. PLoS One, 2012, 7: e34231[本文引用:3][JCR: 3.73]
[16]
KomiyaR, YokoiS, ShimamotoK. A gene network for long-day flowering activates RFT1 encoding a mobile flowering signal in rice. Development, 2009, 136: 3443-3450[本文引用:2][JCR: 6.208]
[17]
Kim SL, LeeS, Kim HJ, Nam HG, AnG. OsMADS51 is a short-day flowering promoter that functions upstream of Ehd1, OsMADS14, and Hd3a. Plant Physiol, 2007, 145: 1484-1494[本文引用:2][JCR: 6.555]
[18]
LeeS, KimJ, Han JJ, Han MJ, AnG. Functional analyses of the flowering time gene OsMADS50, the putative SUPPRESSOR OF OVEREXPRESSION OF CO 1/AGAMOUS-LIKE 20 (SOC1/ AGL20) ortholog in rice. Plant J, 2004, 38: 754-764[本文引用:3][JCR: 6.582]
[19]
Xue WY, Xing YZ, Weng XY, ZhaoY, Tang WJ, WangL, Zhou HJ, Yu SB, Xu CG, Li XH, Zhang QF. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat Genet, 2008, 40: 761-767[本文引用:3][JCR: 35.209]
[20]
Wei XJ, Xu JF, Guo HN, JiangL, Chen SH, Yu CY, Zhou ZL, Hu PS, Zhai HQ, Wan JM. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol, 2010, 153: 1747-1758[本文引用:3][JCR: 6.555]
[21]
Yan WH, WangP, Chen HX, Zhou HJ, Li QP, Wang CR, Ding ZH, Zhang YS, Yu SB, Xin YZ, Zhang QF. A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice. Mol Plant, 2011, 4: 319-330[本文引用:3][JCR: 6.126][CJCR: 0.9487]
[22]
HoriK, Ogiso-TanakaE, MatsubaraK, YamanouchiU, EbanaK, YanoM. Hd16, a gene for casein kinase I, is involved in the control of rice flowering time by modulating the day-length response. Plant J, 2013, 76: 36-46[本文引用:3][JCR: 6.582]
[23]
MatsubaraK, Ogiso-TanakaE, HoriK, EbanaK, AndoT, YanoM. Natural variation in Hd17, a homolog of Arabidopsis ELF3 that is involved in rice photoperiodic flowering. Plant Cell Physiol, 2012, 53: 709-716[本文引用:3][JCR: 4.134]
[24]
Yuan QB, SaitoH, OkumotoY, InoueH, NishidaH, TsukiyamaT, TeraishiM, TanisakaT. Identification of a novel gene ef7 conferring an extremely long basic vegetative growth phase in rice. Theor Appl Genet, 2009, 119: 675-684[本文引用:3][JCR: 3.658]
[25]
徐华山, 孙永健, 周红菊, 余四斌. 构建水稻优良恢复系背景的重叠片段代换系及其效应分析. 作物学报, 2007, 33: 979-986Xu HS, Sun YJ, Zhou HJ, Yu SB. Development and characterization of contiguous segment substitution lines with background of an elite restorer line. Acta Agron Sin, 2007, 33: 979-986 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[26]
YeG, Smith KF. Marker-assisted gene pyramiding for inbred line development: basic principles and practical guidelines. J Plant Breed, 2008, 2: 1-10[本文引用:1]
[27]
GurA, ZamirD. Natural variation can lift yield barriers in plant breeding. PLOS Biol, 2004, 2: 1610-1615[本文引用:1][JCR: 12.69]
[28]
曾瑞珍, 施军琼, 黄朝锋, 张泽民, 丁效华, 李文涛, 张桂权. 籼稻背景的单片段代换系群体的构建. 作物学报, 2006, 32: 88-95Zeng RZ, Shi JQ, Huang CF, Zhang ZM, Ding XH, Li WT, Zhang GQ. Development of a series of single segment substitution lines in indica background of rice (Oryza sativa L. ). Acta Agron Sin, 2006, 32: 88-95 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[29]
罗继景, 黄巍, 朱瑞良, 林鸿宣. 栽培稻抗旱性相关性状QTL的定位. 植物生理学通讯, 2005, 41: 260-268Luo JJ, HuangW, Zhu RL, Lin HX. QTL mapping of drought-resistance of cultivated rice. Plant Physiol Commun, 2005, 41: 260-268 (in Chinese with English abstract)[本文引用:1][CJCR: 0.849]
[30]
Li WT, Zeng RZ, Zhang ZM, Zhang GQ. Mapping of S-b locus for F1 pollen sterility in cultivated rice using PCR based markers. Acta Bot Sin, 2002, 44: 463-467[本文引用:1][JCR: 0.599]
[31]
Paterson AH, DamonS, Hewitt JD, ZamirD, Rabinowitch HD, Lincoln SE, Land er ES, Tanksley SD. Mendelian factors underlying quantitative traits in tomato: comparison across species, generations, and environments. Genetics, 1991, 127: 181-197[本文引用:1][JCR: 4.389]
[32]
Lichtenthaler HK. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol, 1987, 48: 350-382[本文引用:1][JCR: 2.002]
PanaudO, ChenX, McCouch S R. Development of microsatellite markers and characterization of simple sequence length polymorphism in rice (Oryza sativa L). Mol Genet Genomics, 1996, 259: 597-607[本文引用:1][JCR: 2.881]
[35]
桑贤春, 何光华, 张毅, 杨正林, 裴炎. 水稻PCR扩增模板的快速制备. 遗传, 2003, 25: 705-707Sang XC, He GH, ZhangY, Yang ZL, PeiY. The simple grain of templates of rice genomes DNA for PCR. Hereditas (Beijing), 2003, 25: 705-707 (in Chinese with English abstract)[本文引用:1]
[36]
郎有忠, 窦永秀, 王美娥, 张祖建, 朱庆森. 水稻生育期对籽粒产量及品质的影响. 作物学报, 2012, 38: 528-534Lang YZ, Dou YX, Wang ME, Zhang ZJ, Zhu QS. Effects of growth duration on grain yield and quality in rice (Oryza sativa L. ). Acta Agron Sin, 2012, 38: 528-534 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[37]
KomiyaR, IkegamiA, TamakiS, YokoiS, ShimamotoK. Hd3a and RFT1 are essential for flowering in rice. Development, 2008, 135: 767-774[本文引用:1][JCR: 6.208]
[38]
Endo-HigashiN, IzawaT. Flowering time genes Heading date 1 and Early heading date 1 together control panicle development in rice. Plant Cell Physiol, 2011, 52: 1083-1094[本文引用:1][JCR: 4.134]
[39]
HayamaR, IzawaT, ShimamotoK. Isolation of rice genes possibly involved in the photoperiodic control of flowering by a fluorescent differential display method. Plant Cell Physiol, 2002, 43: 494-504[本文引用:1][JCR: 4.134]
[40]
OgisoE, TakahashiY, SasakiT, YanoM, IzawaT. The role of casein kinase II in flowering time regulation has diversified during evolution. Plant Physiol, 2010, 152: 808-820[本文引用:1][JCR: 6.555]
[41]
胡文明, 阚海华, 王伟, 徐辰武. 等位基因功能差异的统计遗传学分析及应用. 作物学报, 2014, 40: 72-79Hu WM, Kan HH, WangW, Xu CW. Statistical genetics approach for functional difference identification of allelic variations and its application. Acta Agron Sin, 2014, 40: 72-79 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[42]
姚国新, 李金杰, 张强, 胡广隆, 陈超, 汤波, 张洪亮, 李自超. 利用4个姊妹近等基因群体定位水稻粒重和粒形QTL. 作物学报, 2010, 36: 1310-1317Yao GX, Li JJ, ZhangQ, Hu GL, ChenC, TangB, Zhang HL, Li ZC. QTLs for grain weight and grain shape using 4NILs. Acta Agron Sin, 2010, 36: 1310-1317 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
 , 李燕, 樊亚伟, 许军红, 郑丽媛, 何光华, 杨正林, 王楠, 赵芳明
, 李燕, 樊亚伟, 许军红, 郑丽媛, 何光华, 杨正林, 王楠, 赵芳明









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}