关键词:豌豆; 白粉病; 抗病基因; PsMLO1基因; 功能标记 Identification of Powdery Mildew Resistance Gene in Pea Line X9002 WANG Zhong-Yi1, FU Hai-Ning1,2, SUN Su-Li1, DUAN Can-Xin1, WU Xiao-Fei1, YANG Xiao-Ming2, ZHU Zhen-Dong1,* 1 National Key Facility for Crop Gene Resource and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
2 Gansu Academy of Agricultural Sciences, Lanzhou 730070, China
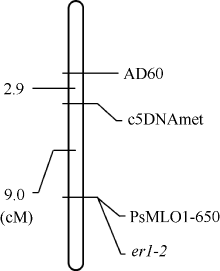
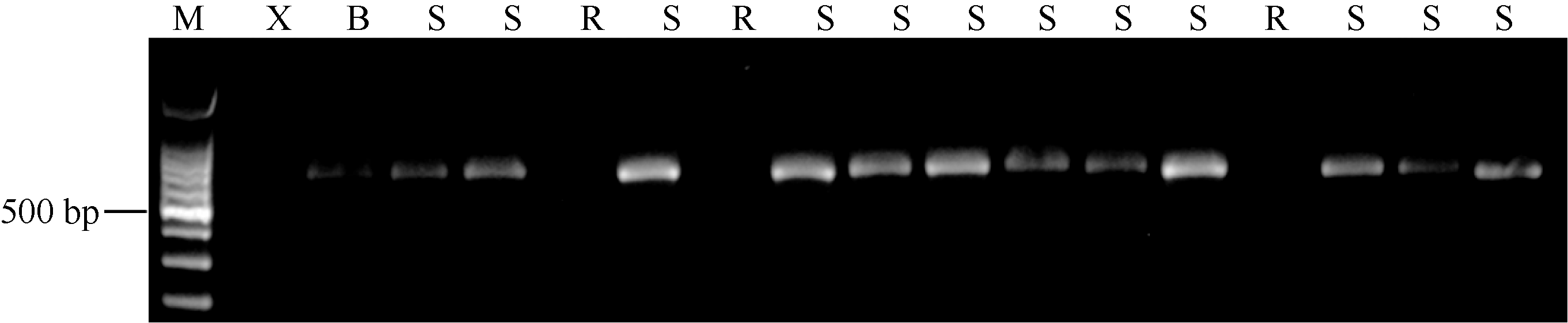
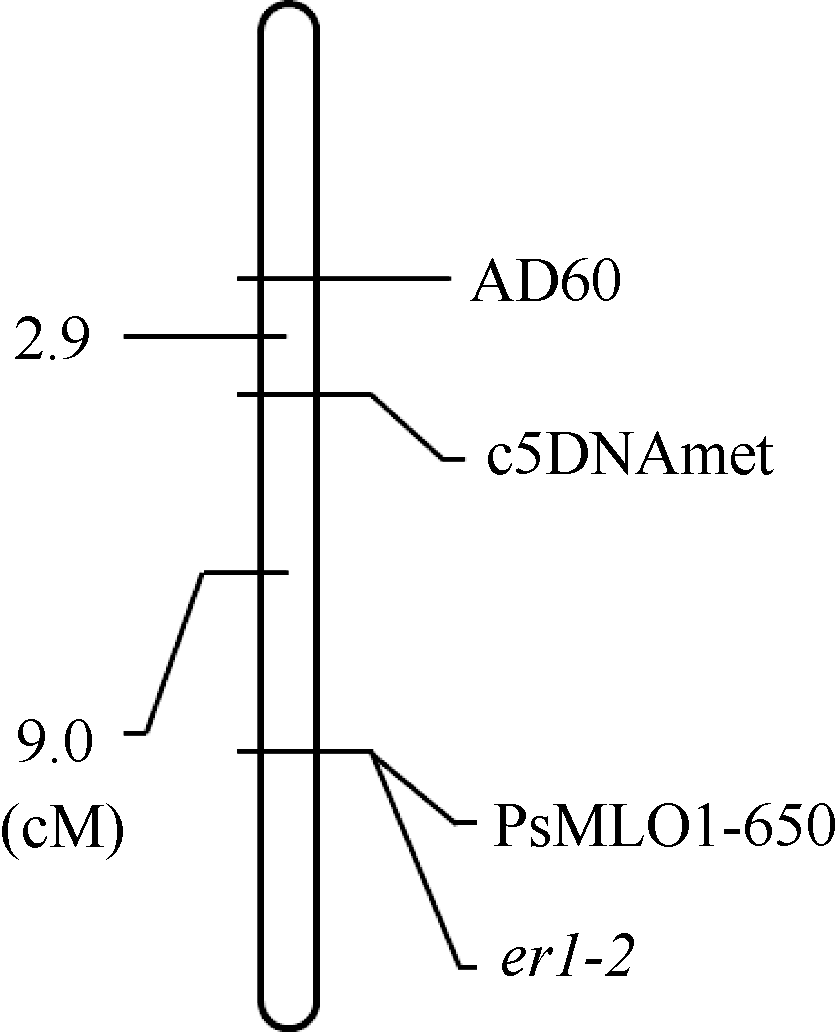
AbstractPowdery mildew is one of the major diseases in pea, causing severe economic loss worldwide. Planting resistant cultivars is the most effective, economical and eco-friendly method for controlling the disease. To date, two recessive resistance genes ( er1, er2) and one dominant resistance gene ( Er3) have been identified in pea, and er1 has been utilized in breeding programs worldwide. Gene er1 is a member of MLOgene family, and er1 resistance is caused by the loss of function at a PsMLO1 locus in pea. X9002 with resistance to powdery mildew is an afilapea line bred by Gansu Academy of Agricultural Sciences. Here, we identified the powdery mildew resistance gene in X9002, and developed molecular marker for the gene selection. Genetic analysis for powdery mildew resistance showed that X9002 carries a recessive resistance gene. The resistance gene was mapped in a region carrying er1 locus on the pea linkage group VI using SSR markers, and was linked to SSR marker AD60 and gene marker c5DNAmet. PsMLO1 sequence analysis revealed that X9002 carries an insertion of unknown size and identity. The same mutation also existed in pea cultivars Stratagem and Franklin carrying er1-2 allele, indicating that the resistance gene is er1-2 in X9002. A functional marker PsMLO1-650 for er1-2 was developed, and the marker was a coupling-phase marker that was detected only in susceptible plants. PsMLO1-650 can be used effectively in marker-assisted selection.
Keyword:Pisum sativumL.; Powdery mildew; Resistance gene; PsMLO1 gene; Functional marker Show Figures Show Figures
图1 部分野生型PsMLO1编码序列与豌豆品系X9002中鉴定的3个PsMLO1 cDNA同源区域的多重比对Fig. 1 Multiple alignment among partial wild-type PsMLO1coding sequence and homologous regions of three PsMLO1 cDNA sequences identified in line X9002
SmýkalP, AubertG, BurstinJ, Coyne CJ, Ellis N T H, Flavell A J, Ford R, Hýbl M, Macas J, Neumann P, McPhee K E, Redden R J, Rubiales D, Weller J L, Warkentin T D. Pea (Pisum sativum L. ) in the genomic era. Agronomy, 2012, 2: 74-115[本文引用:1][JCR: 0.863]
[2]
LiL, Redden RJ, Zong XX, Berger JD, Bennett SJ. Ecogeographic analysis of pea collection sites from China to determine potential sites with abiotic stresses. Genet Resour Crop Evol, 2013, 60: 1801-1815[本文引用:1]
[3]
FAOSTAT2013. Available online: AOSTAT 2013. Available online: http://faostat3.fao.org/(accessed on 13 October 2014[本文引用:1]
[4]
GhafoorA, McPhee K. 2012. Marker assisted selection (MAS) for developing powdery mildew resistant pea cultivars. Euphytica2012, 186: 593-607[本文引用:3][JCR: 1.643]
[5]
FondevillaS, RubialesD. Powdery mildew control in pea: a review. Agron Sust Dev, 2012, 32: 401-409[本文引用:4]
[6]
Harland SC. Inheritance of immunity to mildew in Peruvian forms ofPisum sativum. Heredity, 1948, 2: 263-269[本文引用:3]
[7]
Heringa RJ, NorelA, Tazelaar MF. Resistance to powdery mildew (Erysiphe polygoni DC. ) in peas (Pisum sativum L. ). Euphytica, 1969, 18: 163-169[本文引用:4][JCR: 1.643]
[8]
Tiwari KR, Penner GA, Warkentin TD. Inheritance of powdery mildew resistance in pea. Can J Plant Sci, 1997, 77: 307-310[本文引用:6][JCR: 0.716]
[9]
VaidA, Tyagi PD. Genetics of powdery mildew resistance in pea. Euphytica, 1997, 96: 203-206[本文引用:5][JCR: 1.643]
[10]
SharmaB. The Pisum genus has only one recessive gene for powdery mildew resistance. Pisum Genet, 2003, 35: 22-27[本文引用:3]
[11]
SharmaB. Identification of recessive er gene for powdery mildew resistance in a land race of Pisum sativum. Pisum Genet, 2003, 35: 30-31[本文引用:3]
[12]
Liu SM, O’BrienL, Moore S G. A single recessive gene confers effective resistance to powdery mildew of field pea grown in northern New South Wales. Aust J Exp Agric, 2003, 43: 373-378[本文引用:4][JCR: 1.621]
[13]
FondevillaS, Carver T LW, Moreno MT, RubialesD. Macroscopic and histological characterization of genes er1 and er2 for powdery mildew resistance in pea. Eur J Plant Pathol, 2006, 115: 309-321[本文引用:1][JCR: 1.61]
[14]
FondevillaS, Torres AM, MorenoM T, Rubiales D. Identification of a new gene for resistance to powdery mildew in Pisum fulvum, a wild relative of pea. Breed Sci, 2007, 57: 181-184[本文引用:2][JCR: 1.04]
[15]
DirlewangerE, Isaac PG, RanadeS, BelajouzaM, CousinR, VienneD. Restriction fragment length polymorphism analysis of loci associated with disease resistance genes and developmental traits in Pisum sativum L. Theor Appl Genet, 1994, 88: 17-27[本文引用:2][JCR: 3.658]
[16]
KatochV, SharmaS, PathaniaS, Banayal DK, Sharma SK, RathourR. Molecular mapping of pea powdery mildew resistance gene er2 to pea linkage group III. Mol Breed, 2010, 25: 229-237[本文引用:1][JCR: 3.251]
[17]
Timmerman GM, Frew TJ, Weeden NF. Linkage analysis of er1, a recessive Pisum sativum gene for resistance to powdery mildew fungus (Erysiphe pisi DC. ). Theor Appl Genet, 1994, 88: 1050-1055[本文引用:2][JCR: 3.658]
[18]
Tiwari KR, Penner GA, Warkentin TD. Identification of coupling and repulsion phase RAPD markers for powdery mildew resistance gene er-1 in pea. Genome, 1998, 41: 440-444[本文引用:3][JCR: 1.668]
[19]
JanilaP, SharmaB. RAPD and SCAR markers for powdery mildew resistance gene er1 in pea. Plant Breed, 2004, 12: 271-274[本文引用:2][JCR: 1.175]
[20]
EkM, EklundM, Von PostR, DaytegC, HenrikssonT, WeibullP, CeplitisA, IsaacP, TuvessonS. Microsatellite markers for powdery mildew resistance in pea (Pisum sativum L. ). Hereditas, 2005, 142: 86-91[本文引用:4][CJCR: 0.928]
[21]
PereiraG, MarquesC, RibeiroR, FormigaS, DâmasoM, Sousa TM, FarinhóM, Leitão JM. Identification of DNA markers linked to an induced mutated gene conferring resistance to powdery mildew in pea (Pisum sativum L. ). Euphytica, 2010, 171: 327-335[本文引用:2][JCR: 1.643]
[22]
Srivastava RK, Mishra SK, SinghK, MohapatraT. Development of a coupling-phase SCAR marker linked to the powdery mildew resistance gene er1 in pea (Pisum sativum L. ). Euphytica, 2012, 186: 855-866[本文引用:2][JCR: 1.643]
[23]
Jørgensen JH. Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica, 1992, 63: 141-152[本文引用:1][JCR: 1.643]
[24]
PereiraG, LeitãoJ. Two powdery mildew resistance mutations induced by ENU in Pisum sativum L. affect the locus er1. Euphytica, 2010, 171: 345-354[本文引用:1][JCR: 1.643]
[25]
HumphryM, ReinstädlerA, IvanovS, BisselingT, PanstrugaR. Durable broad-spectrum powdery mildew resistance in pea er1 plants is conferred by natural loss-of-function mutations in PsMLO1. Mol Plant Pathol, 2011, 12: 866-878[本文引用:6][JCR: 3.877]
[26]
PavanS, SchiavulliA, AppianoM, Marcotrigiano AR, CilloF, Visser R GF, BaiY, LottiC, Luigi Ricciardi L. Pea powdery mildew er1 resistance is associated to loss-of-function mutations at a MLO homologous locus. Theor Appl Genet, 2011, 123: 1425-1431[本文引用:4][JCR: 3.658]
[27]
SantoT, RashkovaM, AlabacaC, LeitaoJ. The ENU-induced powdery mildew resistant mutant pea (Pisum sativum L. ) lines S(er1mut1) and F(er1mut2) harbour early stop codons in the PsMLO1 gene. Mol Breed, 2013, 32: 723-727[本文引用:2][JCR: 3.251]
[28]
PavanS, SchiavulliA, AppianoM, MiacolaC, Visser R G F, Bai Y L, Lotti C, Ricciardi L. Identification of a complete set of functional markers for the selection of er1 powdery mildew resistance in Pisum sativum L. Mol Breed, 2013, 31: 247-253[本文引用:3][JCR: 3.251]
[29]
OndřejM, DostálováR, HýblM, OdstrčilováL, TyllerR, TrojanR. Utilization of afila types of pea (Pisum sativum L. ) resistant to powdery mildew (Erysiphe pisi DC. ) in the breeding programs. Plant Soil Environ, 2003, 49: 481-485[本文引用:2][JCR: 1.113]
[30]
曾亮, 李敏权, 杨晓明. 豌豆种质资源白粉病抗性鉴定. 草原与草坪, 2012, 32: 35-38ZengL, Li MQ, Yang XM. Identification of resistance of peas resources to powdery mildew. Grassland & Turf, 2012, 32(4): 35-38 (in Chinese with English abstract)[本文引用:2]
[31]
王仲怡, 包世英, 段灿星, 宗绪晓, 朱振东. 豌豆抗白粉病资源筛选及分子鉴定. 作物学报, 2013, 39: 1030-1038Wang ZY, Bao SY, Duan CX, Zong XX, Zhu ZD. Screening and molecular identification of resistance to powdery mildew in pea germplasm. Acta Agron Sin, 2013, 39: 1030-1038 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[32]
付海宁, 孙素丽, 朱振东, 段灿星, 杨晓明. 加拿大豌豆品种(系)抗白粉病表型和基因型鉴定. 植物遗传资源学报, 2014, 15: 1028-1033Fu HN, Sun SL, Zhu ZD, Duan CX, Yang XM. Phenotypic and genotypic identification of powdery mildew resistance in pea cultivars or lines from Canada. J Plant Genet Resour, 2014, 15: 1028-1033 (in Chinese with English abstract)[本文引用:1][CJCR: 1.1628]
[33]
Rana JC, Banyal DK, Sharma KD, Sharma MK, Gupta SK, Yadav SK. Screening of pea germplasm for resistance to powdery mildew. Euphytica, 2013, 189: 271-282[本文引用:1][JCR: 1.643]
[34]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating population. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1][JCR: 9.737]
[35]
LoridonK, McPhee K, Morin J, Dubreuil P, Pilet-Nayel M L, Aubert G, Rameau C, Baranger A, Coyne C, Lejeune-Henaut I, Burstin J. Microsatellite marker polymorphism and mapping in pea (Pisum sativum L. ). Theor Appl Genet, 2005, 111: 1022-1031[本文引用:2][JCR: 3.658]
[36]
BordatA, SavoisV, NicolasM, SalseJ, ChauveauA, BourgeoisM, PotierJ, HoutinH, RondC, MuratF, MargetP, AubertG, BurstinJ. Translational genomics in legumes allowed placing in silico 5460 unigenes on the pea functional map and identified cand idate genes in Pisum sativum L. Genes Genome Genet, 2011, 1: 93-103[本文引用:1]
[37]
Land er ES, GreenP, AbrahamsonJ, BarlowA, Daly MJ, Lincoln SE, NewburgL. MAPMAKER: an interactive computer package for constructing primary genetic maps of experimental and natural populations. Genomics, 1987, 1: 174-181[本文引用:1][JCR: 3.01]
[38]
Kosambi DD. The estimation of map distances from recombination values. Ann Eugen, 1944, 12: 172-175[本文引用:1]
[39]
SharmaB, YadavY. Pisum fulvum carries a recessive gene for powdery mildew resistance. Pisum Genet, 2003, 35: 31[本文引用:1]
[40]
Pierce WH. Resistance to powdery mildew in peas. Phytopathology, 1948, 38: 21[本文引用:1][JCR: 2.968]
[41]
CousinR. Resistance to powdery mildew in pea. Ann Amélior Plantes, 1965, 15: 93-97[本文引用:1]
[42]
王凤宝, 董立峰, 付金锋, 郑桂茹, 张柏昌, 梁润波, 聂亚琴, 宗云生, 李守训. 超高产豌豆新品种引种及配套栽培技术研究. 河北农业技术师范学院学报, 1998, 12(4): 17-22Wang FB, Dong LF, Fu JF, Zheng GR, Zhang BC, Liang RP, Nie YQ, Zong YS, Li SX. Studies on introduction and culture techniques of a new super high yielding pea variety. J Hebei Agrotech Teachers College, 1998, 12(4): 17-22 (in Chinese with English abstract)[本文引用:1]
[43]
董立峰, 王凤宝, 付金锋. 半无叶型甜豌豆新品种须菜1号的选育. 长江蔬菜, 2008, (8): 45-46Dong LF, Wang FB, Fu JF. Development of a new semi-leafless sweet pea “Xucai No. 1”. J Changjiang Vegetables, 2008, (8): 45-46 (in Chinese)[本文引用:1][CJCR: 0.2667]
[44]
Marx GA. Location of er proving elusive. Pisum Newsl, 1986, 18: 39-41[本文引用:1]





{kind=link}
{kind=link}
{kind=link}