关键词:拟南芥; 下胚轴向光弯曲; 蓝光; 图位克隆 Functional Analysis and Mapping of Gene P2SA2 Involved in Hypocotyl Phototropism of Arabidopsis thaliana ZHAO Qing-Ping, ZHAO Xiang, MU Shi-Chao, XIAO Hui-Li, ZHANG Xiao* State Key Laboratory of Cotton Biology / Key Laboratory of Plant Stress Biology / College of Life Sciences, Henan University, Kaifeng 475004, China
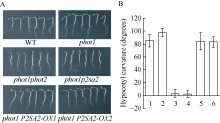
AbstractPHOT1 functions at both low- and high-intensity blue light to mediate phototropic responses, but PHOT2 functions only at high-intensity blue light. The functional redundancy of PHOT1 and PHOT2 on high-intensity blue light-induced phototropic curvature of hypocotyls in Arabidopsis thaliana, restricts the understanding of the mechanism of PHOT2 signal transduction. Therefore, in order to avoid the interference of PHOT1, Arabidopsis phot1 mutant was selected as material for screening high blue light insensitive mutants by the EMS mutation, and we successfully screened and cloned the gene P2SA2 ( phototropin 2 signaling associated 2). The gene P2SA2turned out to be the allelic of NPH3(Nonphototropic hypocotyl 3). The mutation of gene P2SA2 could result in that Arabidopsis thaliana lost the phototropism to unilateral high intensity blue light. Transgenic plants of p2sa2 35S::P2SA2 restored hypocotyl phototropism to high intensity blue light. These findings will open new perspectives about the screening and functional identification of PHOT2 downstream genes in response to high blue light, and provide the theoretical basis to uncover hypocotyl bending mechanism regulated by PHOT2.
Keyword:Arabidopsis thaliana; Blue light; Phototropism; Map-based cloning Show Figures Show Figures
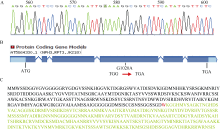
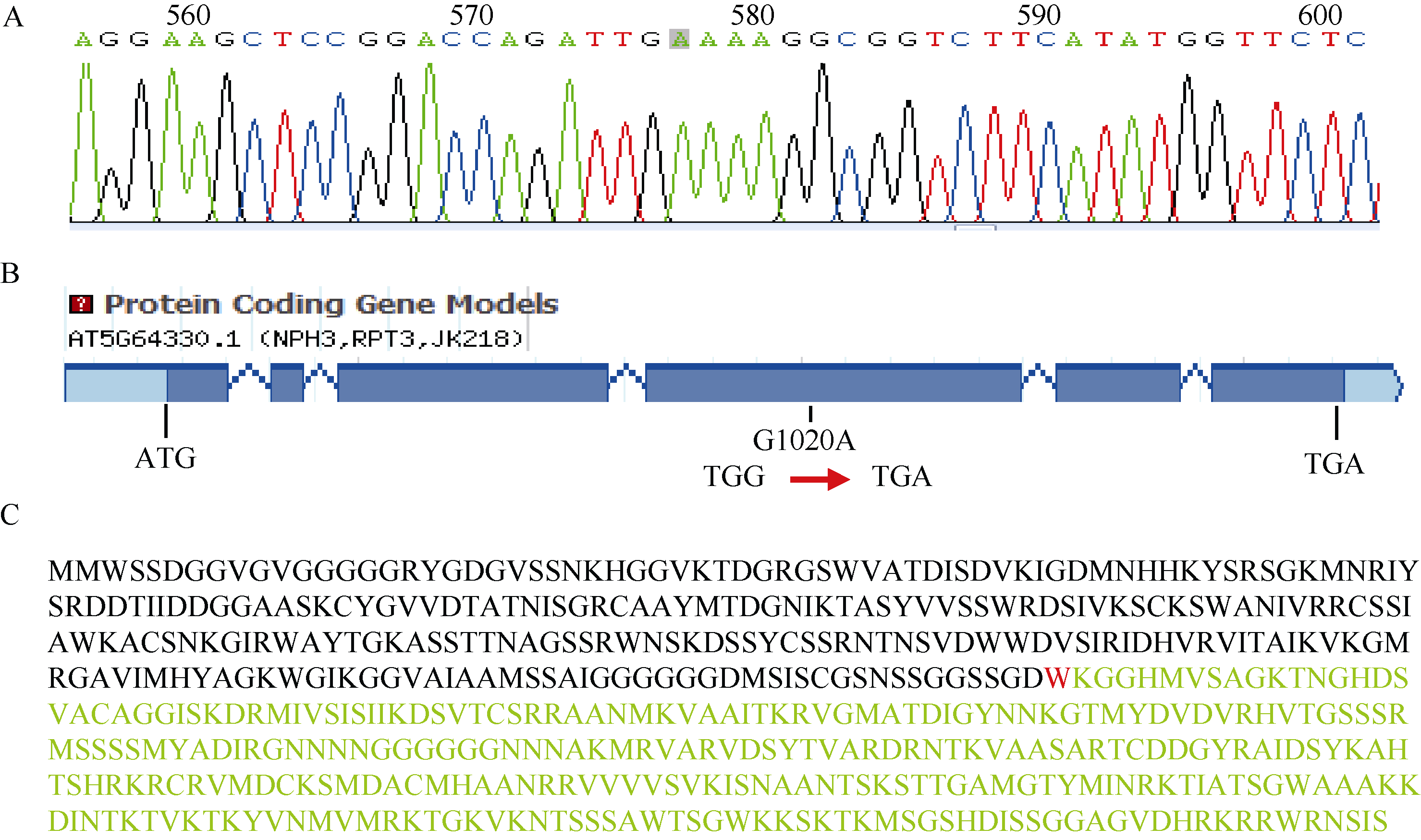
图2P2SA2的定位及测序突变位点分析 A: P2SA2的突变位点; B: p2sa2突变体的突变分析; C: P2SA2基因编码蛋白序列及突变分析, 编码第255位色氨酸(红色W)密码子突变为终止密码子。Fig. 2 Map-based cloning of P2SA2gene and sequence of the mutant gene A: mutation of P2SA2 gene; B: mutation analysis of p2sa2; C: P2SA2 protein sequence. The 255th codon which encodes tryptophan (red W) was mutated to the terminator.
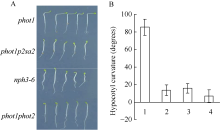
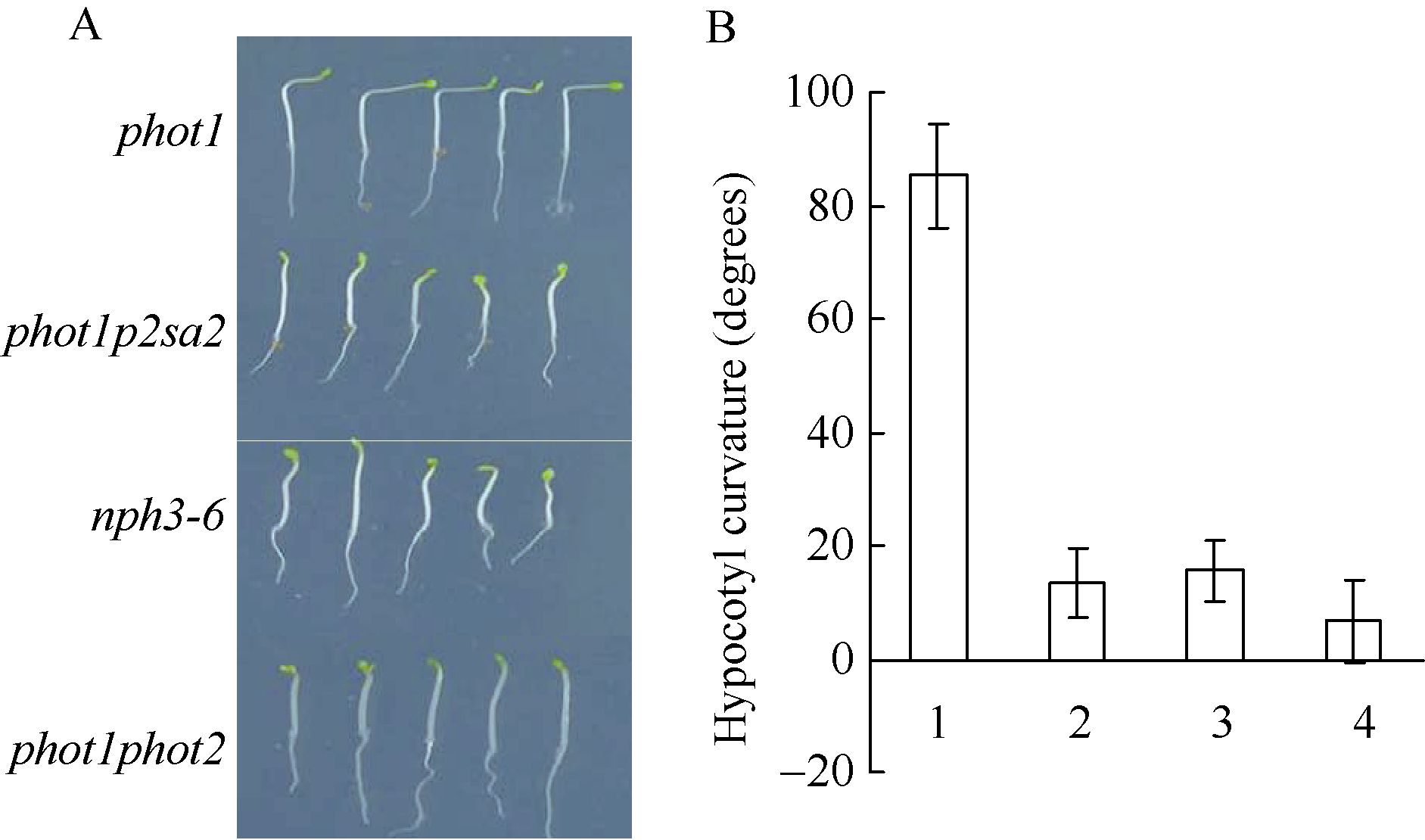
图3 NPH3调节强蓝光诱导的下胚轴弯曲度生长 A: 100 µ mol m-2 s-1蓝光单侧处理12 h后不同基因型拟南芥下胚轴向光弯曲表型; B: 不同基因型拟南芥下胚轴向光弯曲度统计。图中每个数据分别来自3次独立重复试验, 大约19~21颗苗的平均值± 标准误。其中1: phot1; 2: phot1p2sa2; 3:nph3-6; 4: phot1phot2。Fig. 3 Development of phototropic curvature in etiolated seedlings of indicated lines A: development of phototropic curvature in etiolated indicated lines at 100 µ mol m-2 s-1 blue light for 12 hours; B: hypocotyl curvatures were measured in indicated lines. 1:phot1; 2:phot1p2sa2; 3:nph3-6; 4:phot1phot2. Values are the means ± SD (n = 19-21).
HualaE, Oeller PW, LiscumE, Han IS, LarsenE, Briggs WR. Arabidopsis NPH1: a protein kinase with a putative redox- sensing domain. Science, 1997, 278: 2120-2123[本文引用:1]
[2]
DemarsyE, FankhauserC. Higher plants use LOV to perceive blue light. Curr Opin Plant Biol, 2009, 12: 69-74[本文引用:1][JCR: 8.455]
[3]
Christie JM. Phototropin blue-light receptors. Annu Rev Plant Biol, 2007, 58: 21-45[本文引用:1][JCR: 25.962]
[4]
SakaiT, KagawaT, KasaharaM, Swartz TE, Christie JM, Briggs WR, WadaM, OkadaK. Arabidopsis nph1 and npl1: blue light receptors that mediate both phototropism and chloroplast relocation. Proc Natl Acad Sci USA, 2001, 98: 6969-6974[本文引用:2][JCR: 9.737]
[5]
KinoshitaT, DoiM, SuetsuguN, KagawaT, WadaM, Shimazaki K. phot1 and phot2 mediate blue light regulation of stomatal opening. Nature, 2001, 414: 656-660[本文引用:1][JCR: 38.597]
Carbonnel MD, DavisP, Roelfsema MR, InoueS, SchepensI, LariguetP, GeislerM, ShimazakiK, HangarterR, FankhauserC. The Arabidopsis PHYTOCHROME KINASE SUBSTRATE2 protein is a phototropin signaling element that regulates leaf flattening and leaf positioning. Plant Physiol, 2010, 152: 1391-1405[本文引用:1][JCR: 6.555]
[8]
KasaharaM, KagawaT, OikawaK, SuetsuguN, MiyaoM, WadaM. Chloroplast avoidance movement reduces photodamage in plant. Nature, 2002, 420: 829-832[本文引用:1]
[9]
TakemiyaA, InoueS, DoiM, KinoshitaT, ShimazakiK. Phototropins promote plant growth in response to blue light in low light environments. Plant Cell, 2005, 17: 1120-1127[本文引用:1][JCR: 9.251]
[10]
Briggs WR, Christie JM. Phototropins 1 and 2: Versatile plant blue-light receptors. Trends Plant Sci, 2002, 7: 204-210[本文引用:1][JCR: 11.808]
[11]
MotchoulskiA, LiscumE. Arabidopsis NPH3: a NPH1 photoreceptor-interacting protein essential for phototropism. Science, 1999, 286: 961-964[本文引用:4]
[12]
InadaS, OhgishiM, MayamaT, OkadaK, SakaiT. RPT2 is a signal transducer involved in phototropic response and stomatal opening by association with phototropin 1 in Arabidopsis thaliana. Plant Cell, 2004, 16: 887-896[本文引用:3][JCR: 9.251]
[13]
LariguetP, SchepensI, HodgsonD, Pedmale UV, TrevisanM, Kami C, de Carbonnel M, Alonso J M, Ecker J R, Liscum E, Fankhauser C. PHYTOCHROME KINASE SUBSTRATE 1 is a phototropin1 binding protein required for phototropism. Proc Natl Acad Sci USA, 2006, 103: 10134-10139[本文引用:3][JCR: 9.737]
[14]
Blakeslee JJ, Band yopadhyayA, Peer WA, Makam SN, Murphy AS. Relocalization of the PIN1 auxin efflux facilitator plays a role in phototropic responses. Plant Physiol, 2004, 134: 28-31[本文引用:2][JCR: 6.555]
[15]
Stone BB, Stowe-Evans E L, Harper R M, Celaya R B, Ljung K, Sand berg G, Liscum E. Distruption in AUX1-dependent auxin influx alter hypocotyl phototropism inArabidopsis. Mol Plant, 2008, 1: 129-144[本文引用:2]
[16]
SakaiT, WadaT, IshiguroS, OkadaK. RPT2: a signal transducer of the phototropic response in Arabidopsis. Plant Cell, 2000, 12: 225-236[本文引用:1][JCR: 9.251]
[17]
Tseng TS, Briggs WR. The Arabidopsis rcn1-1 mutation impairs dephosphorylation of phot2, resulting in enhanced blue light responses. Plant Cell, 2010, 22: 392-402[本文引用:1][JCR: 9.251]
[18]
DoiM, ShigenagaA, EmiT, KinoshitaT, Shimazaki KI. A transgene encoding a blue-light receptor, phot1, restores blue- light responses in the Arabidopsis phot1phot2 double mutant. J Exp Bot, 2004, 396: 517-523[本文引用:1][JCR: 5.242]
[19]
赵翔, 王琳丹, 李园园, 赵青平, 张骁. PHOT2介导拟南芥下胚轴向光弯曲调节子的筛选与鉴定. 植物学报, 2014, 49: 254-261ZhaoX, Wang LD, Li YY, Zhao QP, ZhangX. Isolation and characterization of regulators involved in PHOT2-mediated phototropismof hypocotyls in Arabidopsis. Chin Bull Bot, 2014, 49: 254-261 (in Chinese with English abstract)[本文引用:1]
[20]
ZhaoX, Wang YL, Qiao XR, WangJ, Wang LD, Xu CS, ZhangX. Phototropins function in high-intensity blue light-induced hypocotyl phototropism in Arabidopsis by altering cytosolic calcium. Plant Physiol, 2013, 162: 1539-1551[本文引用:1][JCR: 6.555]
[21]
Briggs WR, Beck CF, Cashmore AR. The phototropin family of photoreceptors. Plant Cell, 2001, 13: 993-997[本文引用:1][JCR: 9.251]
[22]
Pedmale UV, LiscumE. Regulation of phototropic signaling in Arabidopsis via phosphorylation state changes in the phototropin 1-interacting protein NPH3. J Biol Chem, 2007, 282: 19992-20001[本文引用:2][JCR: 4.651]
[23]
Harper RM, Stowe-Evans E L, Luesse D R, Muto H, Tatematsu K, Watahiki M K, Yamamoto K, Liscum E. The NPH4 locus encodes the auxin response factor ARF7, a conditional regulator of differential growth in aerial Arabidopsis tissue. Plant Cell, 2000, 12: 757-770[本文引用:1][JCR: 9.251]
[24]
LiscumE, Briggs WR. Mutations of Arabidopsis in potential transduction and response components of the phototropic signaling pathway. Plant Physiol, 1996, 112: 291-296[本文引用:1][JCR: 6.555]
[25]
de CarbonnelM, DavisP, Roelfsema MR, InoueS, SchepensI, LariguetP, GeislerM, ShimazakiK, HangarterR, FankhauserC. The Arabidopsis PHYTOCHROME KINASE SUBSTRATE2 protein is a phototropin signaling element that regulates leaf flattening and leaf positioning. Plant Physiol, 2010, 152: 1391-1405[本文引用:1][JCR: 6.555]
[26]
Tsuchida-MayamaT, NakanoM, UeharaY, SanoM, FujisawaN, OkadaK, SakaiT. Mapping of the phosphorylation sites on the phototropic signal transducer, NPH3. Plant Sci, 2008, 174: 626-633[本文引用:1][JCR: 2.922]
[27]
KnauerT, DümmerM, Land grafF, ForreiterC. A negative effector of blue light-induced and gravitropic bending in Arabidopsis. Plant Physiol, 2011, 156: 439-447[本文引用:1][JCR: 6.555]
[28]
HaradaA, ShimazakiK. Phototropins and blue light-dependent calcium signaling in higher plants. Photochem Photobiol, 2007, 83: 102-111[本文引用:1][JCR: 2.287]




{kind=link}
{kind=link}
{kind=link}
{kind=link}