关键词:磷脂酰乙醇胺结合蛋白; 亚洲棉; 雷蒙德氏棉; 成花素; 基因表达 Identification of PEBP Gene Family in Gossypium arboreum and Gossypium raimondii and Expression Analysis of the Gene Family in Gossypium hirsutum LI Chao1, ZHANG Yan-Nan1, LIU Huan-Long1, HUANG Xian-Zhong1,2,* 1 Key Laboratory of Agrobiotechnology / College of Life Sciences, Shihezi University, Shihezi 832003, China
2 Plant Genome Mapping Laboratory, University of Georgia, Athens 30605, GA, USA
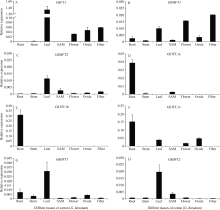
AbstractThe phosphatidylethanolamine-binding proteins (PEBP) widely exist in eukaryotes. In angiosperms, PEBP family genes play important role in promoting or inhibiting flowering, as well as plant architecture control. Eight PEBP genes were identified from diploid cotton Gossypium arboreum (A2) and Gossypium raimondii (D5) genome database, respectively . All the PEBP genes of cotton contained four exons and three introns, and their encoded proteins contained a conserved PEBP motif and critical amino acid sites of PEBP family, which indicated there were at least eight PEBP genes in diploid cotton. Phylogenetic analysis showed that eight cotton PEBP genes comprised three subfamilies: FLOWERING LOCUS T ( FT)-like containing one gene, TERMINAL FLOWER 1 ( TFL1)-like containing five genes including three TFL1 and two BFT genes, and MOTHER OF FT AND TFL1 ( MFT)-like containing two genes. The expression patterns of eight Gossypium hirsutum PEBP family genes in root, stem, leaf, shoot apical meristem, flower, ovule and 25 days post-anthesis (DPA) fiber were determined with quantitative Real-time reverse transcription PCR (qRT-PCR). The results showed that FT1 transcript was preferentially expressed in leaf and secondly in fiber, ovule and flower. MFT1 expressed in all the tissues, with the highest expression level in fiber, then in flower and leaf, while MFT2 transcript was preferentially expressed in leaf. TFL1a, TFL1b, and TFL1c expressed mainly in root, and TFL1c also expressed in leaf, flower and ovule. Expression of BFT1 and BFT2 were present mainly in leaf, and that of BFT1 in the other six tissues except for shoot apical meristem (SAM) were higher than that of BFT2. Expression analysis revealed that eight PEBP genes in cotton have different expression patterns, showing their different functional roles in regulation of cotton development.
Keyword:PEBP; Gossypium arboreum; Gossypium raimondii; Florigen; Gene expression Show Figures Show Figures
表2 雷蒙德氏棉(G. raimondii, D5)基因组中的PEBP家族基因 Table 2 PEBP family genes in G. raimondii (D5) genome
D5基因组转录名 Transcript name of D5 genome
染色体定位 Chromosome in D5 genome
PEBP家族基因 PEBP family gene
开放阅读框/基因组a Open reading frame /genome (bp)a
棉花中同源基因b Homologous gene of cotton b
Gorai.004G264600.1
4
GrFT1
525/3145
GhFT1(HM631972.1)
Gorai.009G174600.2
9
GrMFT1
528/902
GbMFT1(KC513744.1)
Gorai.006G192300.1
6
GrMFT2
519/1872
GbMFT2(KF739071.1)
Gorai.001G121800.1
1
GrTFL1a
525/1022
GrTFL1a(EU026438.1)
Gorai.006G155800.1
6
GrTFL1b
519/1078
GrTFL1b(EU026439.1)
Gorai.009G403800.1
9
GrTFL1c
510/1393
Gorai.004G120400.1
4
GrBFT1
525/796
Gorai.007G010800.1
7
GrBFT2
522/784
a Genome length refers to the length of the corresponding genomic region of ORF (bp); b The contents in parentheses represent the GenBank accession number. a基因组长度指与ORF相对应基因组区域的长度(bp); b括号内为GenBank登录号。
表2 雷蒙德氏棉(G. raimondii, D5)基因组中的PEBP家族基因 Table 2 PEBP family genes in G. raimondii (D5) genome
表3 Table 3 表3(Table 3)
表3 亚洲棉(G. arboreum, A2)基因组中的PEBP家族基因 Table 3 PEBP family genes in G. arboreum (A2) genome
GenBank登录号 GenBank accession No.
E值 E-value
染色体定位 Chromosome in A2 genome
PEBP家族基因 PEBP family gene
开放阅读框/基因组a Open reading frame/ genome (bp)a
棉花中同源基因b Homologous gene of cottonb
AYOE01007731.1
3E-75
3
GaFT1
525/3131
GhFT1(HM631972.1)
AYOE01027581.1
3E-45
10
GaMFT1
528/890
GbMFT1(KC513744.1)
AYOE01031795.1
1E-29
11
GaMFT2
519/1874
GbMFT2(KF739071.1)
AYOE01002909.1
2E-34
1
GaTFL1a
525/1002
GaTFL1a(EU026434.1)
AYOE01032160.1
6E-43
11
GaTFL1b
519/1091
GaTFL1b(EU026442.1)
AYOE01036587.1
1E-35
12
GaTFL1c
510/1358
AYOE01008523.1
1E-40
3
GaBFT1
525/794
AYOE01031581.1
2E-26
11
GaBFT2
522/792
aGenome length refers to the length of the corresponding genomic region of ORF (base pairs). b The contents in parentheses represent the GenBank accession number. a基因组长度指与ORF相对应基因组区域的长度(bp); b括号内为GenBank登录号。
表3 亚洲棉(G. arboreum, A2)基因组中的PEBP家族基因 Table 3 PEBP family genes in G. arboreum (A2) genome
表4 Table 4 表4(Table 4)
表4 棉花PEBP基因家族EST序列信息 Table 4 EST sequences of PEBP family genes in cotton
GenBank登录号 GenBank accession No.
E值 E-value
来源 Origin
基因名称 Gene name
JK805297.1
4E-96
海岛棉Hai7142, 6~24 DPA纤维 G. barbadense Hai7142, 6-24 DPA fiber
GhFT1
ES826802.1
5E-95
陆地棉Xu142, 0~10 DPA纤维 G. hirsutum Xu142, 0-10 DPA fiber
GhFT1
JG852183.1
4E-80
亚洲棉Shixiya, 种子、幼苗、叶、花苞、花和棉铃 G. arboreum Shixiya, seed, seedling, leaf, bud, flower, and boll
GhFT1
ES800631.1
6E-44
陆地棉Xu142, 0~10 DPA纤维 G. hirsutum Xu142, 0-10 DPA fiber
GbMFT1
DW499267.1
2E-33
陆地棉Des119, 顶端分生组织、根和幼嫩纤维 G. hirsutum Des119, SAM, root, young fiber
GbMFT1
BQ410064.1
3E-29
亚洲棉AKA8400, 7~10 DPA纤维 G. arboreum AKA8400, 7-10 DPA fiber
GbMFT1
DW506990.1
6E-46
陆地棉Des119, SAM、根和幼嫩纤维 G. hirsutum Des119, SAM, root, and young fiber
GbMFT2
DW506935.1
1E-34
陆地棉Des119, SAM、根和幼嫩纤维 G. hirsutum Des119, SAM, root, and young fiber
GbMFT2
表4 棉花PEBP基因家族EST序列信息 Table 4 EST sequences of PEBP family genes in cotton
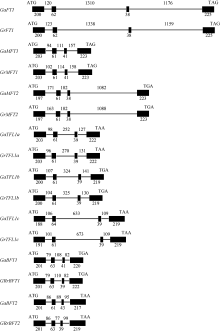
图1 亚洲棉和雷蒙德氏棉PEBP家族基因外显子及内含子分析 ATG和TGA/TAG/TAA分别代表起始密码子和终止密码子, 黑色方框和直线分别代表外显子和内含子, 数字表示外显子和内含子的长度(bp)。Fig. 1 Exons and introns analysis of G. arboreum and G. raimondii PEBP homologs ATG and TGA/TAG/TAA represent initiation codon and termination codon, respectively. Black box and lines represent exonic and intronic regions, respectively. Numbers indicate the length of exons and introns (base pairs).
图2 棉花与拟南芥PEBP家族蛋白序列比对 Consensus pattern: PEBP家族蛋白保守序列; 三角形代表决定FT/TFL1功能的关键氨基酸位点Tyr85 (Y)/His88 (H)和Gln140 (Q)/Asp144 (D)。Fig. 2 Multiple sequence alignment of cotton PBEP homologs and the member of PEBP family in Arabidopsis Consensus pattern: the consensus pattern of PEBP family; Triangle indicates amino acids Tyr85 (Y)/His88 (H) and Gln140 (Q)/Asp144 (D).
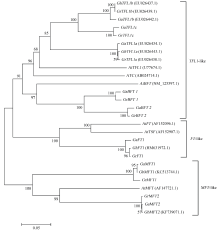
图3 棉花与拟南芥PEBP家族基因进化分析 括号内为GenBank登录号Fig. 3 A Neighbor-Joining phylogenetic analysis of cotton PEBP homologs and the PEBP family in Arabidopsis The GenBank accession number is in parenthese.
Banfield MJ, Barker JJ, Perry AC, Brady RL. Function from structure? The crystal structure of human phosphatidylethanolamine-binding protein suggests a role in membrane signal transduction. Structure, 1998, 6: 1245-1254[本文引用:1][JCR: 5.994]
[2]
HengstU, AlbrechtH, HessD, MonardD. The phosphatidylethanolamine-binding protein is the prototype of a novel family of serine protease inhibitors. J Biol Chem, 2001, 276: 535-540[本文引用:1][JCR: 4.651]
[3]
ChautardH, JacquetM, SchoentgenF, BureaudN, BénédettiH. Tfs1p, a member of the PEBP family, inhibits the Ira2p but not the Ira1p Ras GTPase-activating protein in Saccharomyces cerevisiae. Eukaryot Cell, 2004, 3: 459-470[本文引用:1][JCR: 3.586]
[4]
ChardonF, DamervalC. Phylogenomic analysis of the PEBP gene family in cereals. J Mol Evol, 2005, 61: 579-590[本文引用:1][JCR: 2.145]
[5]
LiQ, FanC, ZhangX, WangX, WuF, HuR, FuY. Identification of a soybean MOTHER OF FT AND TFL1 homolog involved in regulation of seed germination. PLoS One, 2014, 9: e99642[本文引用:4][JCR: 3.73]
[6]
XiW, LiuC, HouX, YuH. MOTHER OF FT AND TFL1 regulates seed germination through a negative feedback loop modulating ABA signaling in Arabidopsis. Plant Cell, 2010, 22: 1733-1748[本文引用:2][JCR: 9.251]
[7]
NakamuraS, AbeF, KawahigashiH, NakazonoK, TagiriA, MatsumotoT, UtsugiS, OgawaT, Hand aH, IshidaH, MoriM, KawauraK, OgiharaY, MiuraH. A wheat homolog of MOTHER OF FT AND TFL1 acts in the regulation of germination. Plant Cell, 2011, 23: 3215-3229[本文引用:3][JCR: 9.251]
[8]
KarlgrenA, Gyllenstrand N, KallmanT, Sundstrom JF, MooreD, LascouxM, LagercrantzU. Evolution of the PEBP gene family in plants: functional diversification in seed plant evolution. Plant Physiol, 2011, 156: 1967-1977[本文引用:6][JCR: 6.555]
[9]
CorbesierL, VincentC, JangS, FornaraF, FanQ, SearleI, GiakountisA, FarronaS, GissotL, TurnbullC, Coupland G. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science, 2007, 316: 1030-1033[本文引用:2]
[10]
TamakiS, MatsuoS, WongH, YokoiS, ShimamotoK. Hd3a protein is a mobile flowering signal in rice. Science, 2007, 316: 1033-1036[本文引用:2]
[11]
LifschitzE, EviatarT, RozmanA, ShalitA, GoldshmidtA, AmsellemZ, Alvarez JP, EshedY. The tomato FT ortholog triggers systemic signals that regulate growth and ?owering and substitute for diverse environmental stimuli. Proc Natl Acad Sci USA, 2006, 103: 6398-6403[本文引用:2][JCR: 9.737]
[12]
AbeM, KobayashiY, YamamotoS, DaimonY, YamaguchiA, IkedaY, IchinokiH, NotaguchiM, GotoK, ArakiT. FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science, 2005, 309: 1052-1056[本文引用:1]
[13]
Lin MK, BelangerH, Lee YJ, Varkonyi-GasicE, TaokaK, MiuraE, Xoconostle-CázaresB, GendlerK, Jorgensen RA, PhinneyB, Lough TJ, Lucas WJ. FLOWRING LOCUS T protein may act as the long-distance florigenic signal in the cucurbits. Plant Cell, 2007, 19: 1488-1506[本文引用:2][JCR: 9.251]
[14]
ImamuraT, NakatsukaT, HiguchiA, NishiharaM, TakahashiH. The gentian orthologs of the FT/TFL1 gene family control floral initiation in Gentiana. Plant Cell Physiol, 2011, 52: 1031-1041[本文引用:2][JCR: 4.134]
[15]
KotodaN, HayashiH, SuzukiM, IgarashiM, HatsuyamaY, KidouS, IgasakiT, NishiguchiM, YanoK, ShimizuT, TakahashiS, IwanamiH, MoriyaS, AbeK. Molecular characterization of FLOWERING LOCUS T-like genes of apple (Malus × domestica Borkh). Plant Cell Physiol, 2011, 51: 561-575[本文引用:2][JCR: 4.134]
[16]
HarigL, Beinecke FA, OltmannsJ, MuthJ, MüllerO, RüpingB, Twyman RM, FischerR, PrüferD, Noll GA. Proteins from the FLOWERING LOCUS T-like subclade of the PEBP family act antagonistically to regulate floral initiation in tobacco. Plant J, 2012, 72: 908-921[本文引用:2][JCR: 6.582]
[17]
Pin PA, BenllochR, BonnetD, Wremerth-WeichE, KraftT, Gielen JJ, NilssonO. An antagonistic pair of FT homologs mediates the control of flowering time in sugar beet. Science, 2010, 330: 1397-1400[本文引用:1]
[18]
BradleyD, RatcliffeO, VincentC, CarpenterR, CoenE. Inflorescence commitment and architecture in Arabidopsis. Science, 1997, 275: 80-83[本文引用:1]
[19]
BradleyD, CarpenterR, CopseyL, VincentC, RothsteinS, CoenE. Control of inflorescence architecture in Antirrhinum. Nature, 1996, 379: 791-797[本文引用:1][JCR: 38.597]
[20]
HanzawaY, MoneyT, BradleyD. A single amino acid converts a repressor to an activator of flowering. Proc Natl Acad Sci USA, 2005, 102: 7748-7753[本文引用:1][JCR: 9.737]
[21]
Ahn JH, MillerD, Winter VJ, Banfield MJ, Lee JH, Yoo SY, Henz SR, Brady RL, WeigelD. A divergent external loop confers antagonistic activity on floral regulators FT and TFL1. EMBO J, 2006, 25: 605-614[本文引用:1][JCR: 9.822]
[22]
WendelJ, BrubakerC, AlvarezI, CronnR. Genetics and Genomics of Cotton. New York: Springer-Verlag, 2009. pp 3-22[本文引用:1]
[23]
东锐, 院海英, 顾超, 郑银英, 黄先忠, 崔百明. 棉花GhFTL1基因的克隆及初步功能分析. 棉花学报, 2011, 23: 515-521DongR, Yuan HY, GuC, Zheng YY, Huang XZ, Cui BM. Clone and primary analysis of the function of GhFTL1 gene in cotton (Gossypium hirsutum). Cotton Sci, 2011, 23: 515-521 (in Chinese with English abstract)[本文引用:5][CJCR: 1.462]
[24]
顾超, 李超, 李晓波, 肖向文, 崔百明, 黄先忠. 海岛棉GbMFT1基因的克隆及表达分析. 作物学报, 2013, 39: 1391-1399GuC, LiC, Li XB, Xiao XW, Cui BM, Huang XZ. Cloning and expression analysis of GbMFT1 gene in Gossypium barbadense L. Acta Agron Sin, 2013, 39: 1391-1399 (in Chinese with English abstract)[本文引用:6][CJCR: 1.667]
[25]
顾超, 郭丹丽, 张峰, 李雪源, 艾先涛, 黄先忠. 海岛棉GbMFT2基因的克隆及表达分析. 棉花学报, 2014, 26: 197-203GuC, Guo DL, ZhangF, Li XY, Ai XT, Huang XZ. Cloning and expression analysis of GbMFT2 gene in Gossypium barbadense L. Cotton Sci, 2014, 26: 197-203 (in Chinese with English abstract)[本文引用:4][CJCR: 1.462]
[26]
ArgiriouA, MichailidisG, Tsaftaris AS. Characterization and expression analysis of TERMINAL FLOWER1 homologs from cultivated alloteraploid cotton (Gossypium hirsutum) and its diploid progenitors. J Plant Physiol, 2008, 165: 1636-1646[本文引用:8][JCR: 2.699]
[27]
Wang KB, Wang ZW, Li FG, Ye WW, Wang JY, Song GL, YueZ, CongL, Shang HH, Zhu SL, Zou CS, LiQ, Yuan YL, Lu CR, Wei HL, Gou CY, Zheng ZQ, YinY, Zhang XY, LiuK, WangB, SongC, ShiN, Kohel RJ, Percy RG, Yu JZ, Zhu YX, WangJ, Yu SX. The draft genome of a diploid cotton Gossypium raimondii. Nat Genet, 2012, 44: 1098-1103[本文引用:1][JCR: 35.209]







{kind=link}
{kind=link}
{kind=link}
{kind=link}