关键词:小麦-中间偃麦草渗入系; 白粉病抗性; GISH; SSR标记; 连锁图谱 Molecular Mapping of Powdery Mildew Resistance Gene PmCH7124 in a Putative Wheat- Thinopyrum intermedium Introgression Line LI Jian-Bo1, QIAO Lin-Yi1,2,*, LI Xin2, ZHANG Xiao-Jun2, ZHAN Hai-Xian2, GUO Hui-Juan2, REN Yong-Kang2, CHANG Zhi-Jian2,* 1Graduate School of Shanxi University, Taiyuan 030006, China
2Institute of Crop Science, Shanxi Academy of Agricultural Sciences / Key Laboratory of Crop Gene Resources and Germplasm Enhancement on Loess Plateau of Ministry of Agriculture, Taiyuan 030031, China
AbstractWheat introgression line CH7124 derived from a cross between wheat- Thinopyrum intermedium line TAI8335 (resistant to powdery mildew) and common wheat variety “Jinmai 33” (susceptible to powdery mildew) exhibits immunity to Blumeria graminisf. sp. tritici ( Bgt) pathotypes E09, E20, E21, E23, E26, Bg1, and Bg2 at the seedling stage. The Bgt resistance in CH7124 is similar to that in TAI8335 and its wild parent Th. intermedium. However, no Th. intermedium chromatin has been detected according to genome in situ hybridization (GISH) assay. In this study, we determined the single dominant Bgt resistance gene in CH7124, tentatively designated PmCH7124, using populations derived from SY95-71/CH7124 (F6) and CH7124/Mianyang 11 (F1 and F2). Five SSR markers ( Xgwm47, Xgwm120, Xwmc332, Xgwm501, and Xbarc101) were identified to be codominant with PmCH7124 according to bulked segregant analysis, and the closely flanking markers were Xgwm501 and Xbacr101with genetic distances of 1.7 cM and 4.5 cM, respectively. The target resistance gene was chromosomally located on 2BL with Chinese Spring nulli-tetrasomicand ditelosomic lines. We primarily consider that PmCH7124is a new Bgt resistance gene because its resistance spectrum, origin, chromosomal location, and linked markers are different from those of the known Bgt resistance genes, such as Pm6, Pm33, PmJM22, MlZec1, MlAB10, and MlLX99.
Keyword:Wheat-Th. intermediumintrogression line; Resistance to powdery mildew; GISH; SSR markers; Linkage map Show Figures Show Figures
表2 小麦CH7124/绵阳11杂交组合不同世代群体对白粉菌菌株E09的成株期抗性分离 Table 2 Segregation of resistance to Bgt pathotypes E09 in different populations derived from the CH7124/Mianyang 11 cross
亲本/组合# Parent/cross#
抗病株 Resistant plants
感病株 Susceptible plants
预期分离比 Expected ratio
P
0
0
1
2
3
合计 Total
5
6
7
8
9
合计 Total
P1 (n = 15)
11
4
15
P2 (n = 15)
2
10
3
15
P3 (n = 15)
1
9
5
15
P2× P1 F1 (n = 15)
6
9
15
P2× P1 F2 (n = 122)
63
19
7
1
1
91
1
3
3
8
16
31
χ 23:1 = 0.01
0.92
P3× P1 F6 (n = 112)
48
7
4
59
2
5
4
15
27
53
χ 21:1 = 0.16
0.57
#P1: CH7124 (resistant); P2: Mianyang 11 (susceptible); P3: SY95-71 (susceptible). n stands for the total plant tested, and no resistant plant with IT=4. #P1: CH7124 (抗病); P2: 绵阳11(感病); P3: SY95-71(感病)。n表示鉴定总株数, 无IT=4的单株。
表2 小麦CH7124/绵阳11杂交组合不同世代群体对白粉菌菌株E09的成株期抗性分离 Table 2 Segregation of resistance to Bgt pathotypes E09 in different populations derived from the CH7124/Mianyang 11 cross
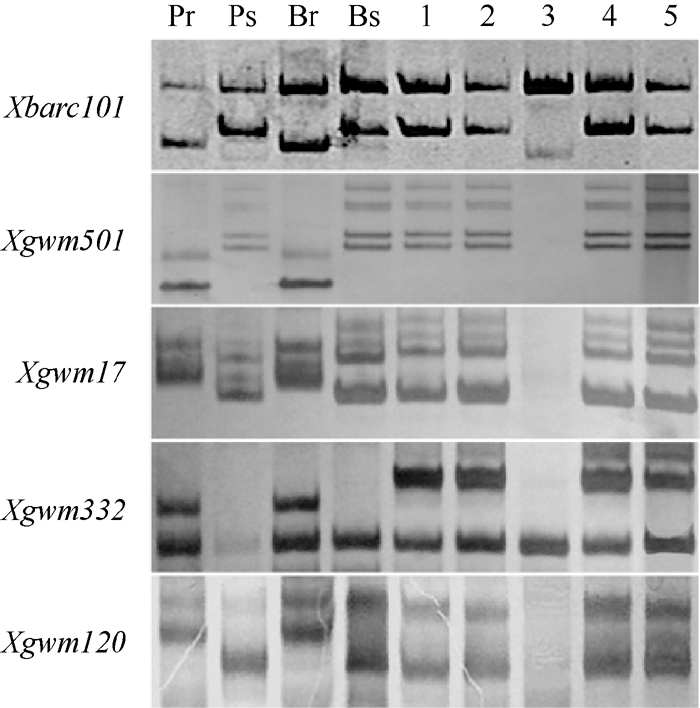
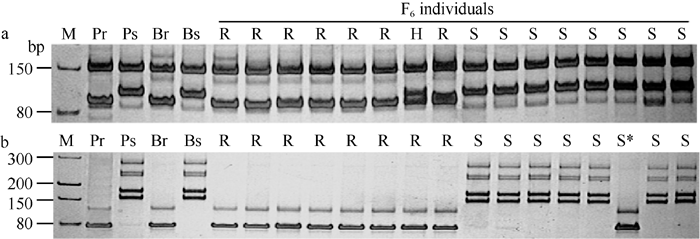
图2PmCH7124侧翼SSR标记Xbarc101 (a)和Xgwm501(b)在SY95-71/CH7124 F6群体中的扩增图谱Fig. 2 Amplification profiles of Xbarc101 (a) and Xgwm501(b), the flanking SSRs of PmCH7124 in SY95-71/CH7124 F6 population
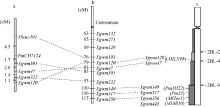
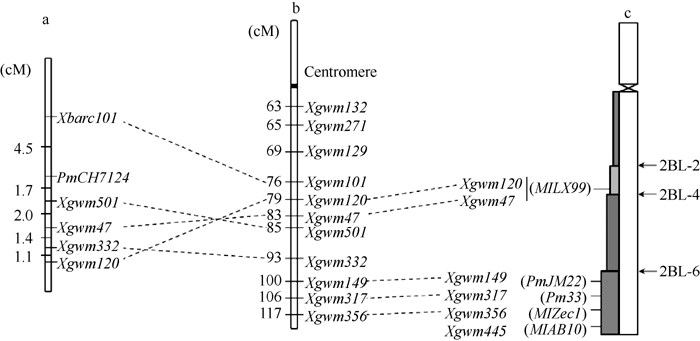
图3PmCH7124的遗传定位(a)及其与已有SSR标记图谱(b)[19]和物理图谱(c)[8]的比较Fig. 3 Genetic mapping of PmCH7124(a) and comparison with genetic map with SSR markers (b) and physical maps (c)
表3 小麦2BL上其他抗病基因连锁标记在SY95-71/CH7124 F6作图群体中的多态性 Table 3 Polymorphism of SSR markers associated with known Bgt resistance genes on chromosome 2BL in the SY95-71/CH7124 F6 mapping population
抗病基因 Resistance gene
连锁标记 Marker
距离 Distance (cM)
亲本 Parent
抗感池 Bulk
PmCH7124
Pm6
XCINAU123
0.1
+
-
-
Pm6
XCINAU127
0.0
+
-
-
Pm33
Xwmc317
1.1
-
-
-
PmJM22
Xwmc149
7.7
-
-
-
MlZec1
Xwmc356
2.0
-
-
-
MlAB10
Xwmc445
7.0
-
-
-
MlLX99
Xcfd73
10.9
-
-
-
MlLX99
Xwmc441
5.7
+
-
-
MlLX99
Xgwm120
2.9
+
+
+
MlLX99
Xgwm47
16.6
+
+
+
MlLX99
Xwmc175
25.3
-
-
-
+: polymorphic or linked; -: non-polymorphic or unlinked. +:存在多态性或连锁; -: 不存在多态性或连锁。
表3 小麦2BL上其他抗病基因连锁标记在SY95-71/CH7124 F6作图群体中的多态性 Table 3 Polymorphism of SSR markers associated with known Bgt resistance genes on chromosome 2BL in the SY95-71/CH7124 F6 mapping population
4 结论小麦-中间偃麦草渗入系新种质CH7124对白粉病菌E09免疫, 成株期抗性表现受1对显性核基因PmCH7124控制。该基因被初步定位在小麦2B染色体长臂上, 位于SSR标记gwm501和barc101之间, 遗传距离分别为1.7 cM和4.5 cM, 并且不同于2BL上已报道的其他白粉病抗病基因。 致谢: 山西省农业科学院植物保护研究所原宗英研究员对抗性鉴定给予大力帮助和指导, 谨致谢忱。 The authors have declared that no competing interests exist.
Line RF, Chen XM. Success in breeding for and managing durable resistance to wheat rusts. Plant Dis, 1995, 79: 1254-1255[本文引用:2][JCR: 2.455]
[2]
MohlerV, BauerC, SchweizerG, KempfH, HartlL. Pm50: a new powdery mildew resistance gene in common wheat derived from cultivated emmer. J Appl Genet, 2013, 54: 259-263[本文引用:1][JCR: 1.847]
[3]
TaoW, LiuD, LiuJ, FengY, ChenP. Genetic mapping of the powdery mildew resistance gene Pm6 in wheat by RFLP analysis. Theor Appl Genet, 2000, 100: 564-568[本文引用:1][JCR: 3.658]
[4]
Zhu ZD, Zhou RH, Kong XY, Dong YC, Jia JZ. Microsatellite markers linked to two powdery mildew resistance gene introgressed from Triticum carthlicum accession PS5 into common wheat. Genome, 2005, 48: 585-590[本文引用:3][JCR: 1.668]
[5]
殷贵鸿, 李根英, 何中虎, 刘建军, 王辉, 夏先春. 小麦新品种济麦22抗白粉病基因的分子标记定位. 作物学报, 2009, 35: 1425-1431Yin GH, Li GY, He ZH, Liu JJ, WangH, Xia XC. Molecular mapping of powdery mildew resistance gene in wheat cultivar Jimai 22. Acta Agron Sin, 2009, 35: 1425-1431 (in Chinese with English abstract)[本文引用:3][CJCR: 1.667]
[6]
MohlerV, Zeller FJ, WenzelG, HsamS L K. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell. ): 9. Gene MlZec1 from Triticum dicoccoides-derived wheat line Zecoi-1. Euphytica, 2005, 142: 161-167[本文引用:3]
Zhao ZH, Sun HG, SongW, LuM, HuangJ, Wu LF, Wang XM, Li HJ. Genetic analysis and detection of the gene MlLX99 on chromosome 2BL conferring resistance to powdery mildew in the wheat cultivar Liangxing 99. Theor Appl Genet, 2013, 126: 3081-3089[本文引用:5][JCR: 3.658]
[9]
Huang XQ, HsamS L K, ZellerF J. Identification of powdery mildew resistance genes in common wheat (Triticum aestivum L. ): IX. Cultivars, land racesand breeding lines grown in China. Plant Breed, 1997, 116: 233-238[本文引用:1][JCR: 1.175]
[10]
Li HJ, Wang XM. Thinopyrumponticum and Th. intermedium: the promising source of resistance to fungal and viral diseases of wheat. J Genet Genomics, 2009, 36: 557-565[本文引用:2][JCR: 2.076][CJCR: 1.323]
[11]
Liu SB, Wang HG. Characterization of a wheat-Thinopyron intermedium substitution line with resistance to powdery mildew. Euphytica, 2005, 143: 229-233[本文引用:1][JCR: 1.643]
[12]
李欣, 张晓军, 张丛卓, 詹海仙, 杨足君, 畅志坚. 兼抗白粉、条锈病小偃麦渗入系 CH7124抗性遗传及细胞学鉴定. 植物遗传资源学报, 2012, 13: 577-582LiX, Zhang XJ, Zhang CZ, Zhan HX, Yang ZJ, Chang ZJ. Cytological characterization and resistance inheritance in alien introgression CH7124 from Thinopyrum intermedium. J Plant Genet Resour, 2012, 13: 577-582 (in Chinese with English abstract)[本文引用:2][CJCR: 1.1628]
[13]
LuoP, LuoH, ChangZ, ZhangH, ZhangM, RenZ. Characterization and chromosomal location of Pm40 in common wheat: a new gene for resistance to powdery mildew derived from Elytrigia intermedium. Theor Appl Genet, 2009, 118: 1059-1064[本文引用:1][JCR: 3.658]
[14]
HeR, ChangZ, YangZ, LiuZ, ZhanH, ZhangX, LiuJ. Inheritance and mapping of a powdery mildew resistance gene Pm43 introgressed from Thinopyrum intermedium into wheat. Theor Appl Genet, 2009, 118: 1173-1180[本文引用:1][JCR: 3.658]
[15]
孙翠花, 侯丽媛, 郭慧娟, 张晓军, 贾举庆, 李欣, 詹海仙, 畅志坚. 小麦-中间偃麦草隐形渗入系抗白粉病基因pmCH83的分子定位. 作物学报, 2013, 39: 2107-2114Sun CH, Hou LY, Guo HJ, Zhang XJ, Jia JQ, LiX, Zhan HX, Chang ZJ. Molecular mapping of powdery mildew resistance gene pmCH83 in a putative wheat-Thinopyrum intermedium cryptic introgression line. Acta Agron Sin, 2013, 39: 2107-2114 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[16]
ChangZ, ZhangX, YangZ, ZhanH, LiX, LiuC, ZhangC. Characterization of a partial wheat-Thinopyrum intermedium amphiploid and its reaction to fungal diseases of wheat. Hereditas, 2010, 147: 304-312[本文引用:2][CJCR: 0.928]
[17]
盛宝钦. 用反应型记载小麦苗期白粉病. 植物保护, 1988, 14(1): 49Sheng BQ. Using infection type records the wheat powdery mildew at seedling stage. Plant Prot, 1988, 14(1): 49 (in Chinese)[本文引用:2]
[18]
盛宝钦, 段霞瑜. 用反应型记载小麦成株白粉病"0~9级法"的改进. 北京农业科学, 1991, 9(1): 37-39Sheng BQ, Duan XY. Modification on the evaluation methods of 0-9 level of powdery mildew infection on wheat. Beijing Agric Sci, 1991, 9(1): 37-39 (in Chinese)[本文引用:1]
[19]
Somers DJ, IsaacP, EdwardsK. A high density microsatellite consensus map for bread wheat (Triticum aestivum L. ). Theor Appl Genet, 2004, 109: 1105-1114[本文引用:1][JCR: 3.658]
[20]
QinB, Cao AZ, Wang HY, Chen TT, FrankM, Liu YY, Ji JH, Liu DJ, Chen PD, Wang XE. Collinearity-based marker mining for the fine mapping of Pm6, a powdery mildew resistance gene in wheat. Theor Appl Genet, 2011, 123: 207-218[本文引用:1][JCR: 3.658]
[21]
Wang ZL, Li LH, He ZH, Duan XY, Zhou YL, Chen XM, LillemoM, Singh RP, WangH, Xia XC. Seedling and adult plant resistance to powdery mildew in Chinese bread wheat cultivars and lines. Plant Dis, 2005, 89: 457-463[本文引用:1][JCR: 2.455]
[22]
Zhang XY, KoulA, PetroskiR, OuelletT, FedakG, Dong YS, WangR R C. Molecular verification and characterization of BYDV-resistant germplasms derived from hybrids of wheat with Thinopyrum ponticum and Th. intermedium. Theor Appl Genet, 1996, 93: 1033-1039[本文引用:2][JCR: 3.658]
[23]
ChenQ, Conner RL, LarocheA, AhmadF. Molecular cytogenetic evidence for a high level of chromosome pairing among different genomes in Triticum aestivum-Thinopyrum intermedium hybrids. Theor Appl Genet, 2001, 102: 847-852[本文引用:1][JCR: 3.658]
[24]
张学勇, 董玉琛, 杨欣明. 小麦与长穗偃麦草、中间偃麦草杂种及其衍生后代的细胞遗传学研究: III. 小麦和偃麦草基因重组的遗传基础浅析. 遗传学报, 1995, 22: 217-222Zhang XY, Dong YC, Yang XM. Cytogenetic research on hybrids of Triticum with both Thinopyrum ponticum and Th. intermedium as well as their derivatives: III. Primary detection of genetic base for introgression of useful genes from the two alien species to wheat. Acta Genet Sin, 1995, 22: 217-222 (in Chinese with English abstract)[本文引用:1]
[25]
Dong YS, Bu XL, Luan YS, He MY, LiuB. Molecular characterization of a cryptic wheat-Thinopyrum intermedium translocation line: evidence for genomic instability in nascent allopolyploid and aneuploid lines. Genet Mol Biol, 2004, 27: 237-241[本文引用:1][JCR: 0.744]
KuraparthyV, ChhunejaP, Dhaliwal HS, KaurS, Bowder RL, Gill BS. Characterization and mapping of cryptic alien introgression from Aegilops geniculata with novel leaf rust and stripe rust resistance genes Lr57 and Yr40 in wheat. Theor Appl Genet, 2007, 114: 1379-1389[本文引用:1][JCR: 3.658]
[28]
宋凤景, 肖明纲, 黄江, 王晓鸣, 朱振东, 武小菲, . 李洪杰. 12个小麦品种(系)白粉病抗性的遗传分析. 作物学报, 2012, 38: 1339-1345Song FJ, Xiao MG, HuangJ, Wang XM, Zhu ZD, Wu XF, Li HJ. Inheritance of resistance to powdery mildew in 12 wheat varieties (lines). Acta Agron Sin, 2012, 38: 1339-1345 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
 , 乔麟轶
, 乔麟轶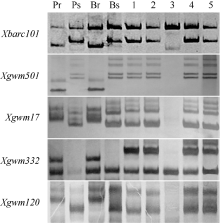

{kind=link}
{kind=link}
{kind=link}