关键词:粗山羊草; 生长素; Aux/IAA基因家族; 染色体定位; 生物信息学 Whole-Genome Sequence Isolation, Chromosome Location, and Characteriza-tion of Primary Auxin-ResponsiveAux/IAA Gene Family inAegilops tauschii QIAO Lin-Yi1,4,**, LI Xin1,**, CHANG Zhi-Jian1, ZHANG Xiao-Jun1, ZHAN Hai-Xian1, GUO Hui-Juan1, LI Jian-Bo4, CHANG Jian-Zhong3,*, ZHENG Jun2,* 1Institute of Crop Science, Shanxi Academy of Agricultural Sciences / Key Laboratory of Crop Gene Resources and Germplasm Enhancement on Loess Plateau, Ministry of Agriculture, Taiyuan 030031, China
2 Wheat Research Institute, Shanxi Academy of Agricultural Sciences, Linfen 041000, China
3 Research Center of Dryland Farming, Shanxi Academy of Agricultural Sciences, Taiyuan 030006, China
4Graduate School of Shanxi University, Taiyuan 030006, China
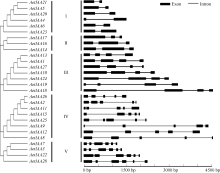
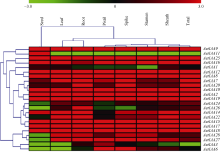
AbstractAuxin, as one of the most important hormones, plays a key role in many processes of plant development. TheAux/IAA family contains important early auxin response genes. A genome-wide research ofAux/IAA genes inAegilops tauschii was carried out using bioinformatic method. In this study, 28Aux/IAA genes were identified inAe. tauschii, which were distributed on seven chromosomes ofAe. tauschii genome. TwentyAux/IAA genes share four conserved amino acid sequence motifs and five were mapped on the same locus with known markers.AetIAA3,AetIAA11, andAetIAA26 were specifically expressed in pistil, seed and root ofAe. tauschii, respectively. Eleven pairs ofAe. tauschii-Triticum urartuand five pairs ofAe. tauschii-Hordeum vulgare Aux/IAA proteins were orthologous in the phylogenetic tree. Collinearity analysis indicatedAux/IAA genes showed a higher synteny betweenAe. tauschii and two other species (Brachypodium distachyon andOryza stativa). TheAux/IAA genes isolated can not only be applied in genetic improvement of common wheat but also provide basic information in further research ofAux/IAA genes in wheat.
Keyword:Aegilops tauschii; Auxin; Aux/IAA gene family; Chromosome location; Bioinformatics Show Figures Show Figures
图1 粗山羊草 Aux/IAA家族基因的染色体定位基因左边的竖线表示这些基因位于同一位点。Fig. 1 Chromosomal distribution of Aux/ IAA genes in Ae. tauschii genomeThe vertical line, which is at the left side of some genes or markers, indicated that these genes or marks are located at the same locus.
表1 Table 1 表1(Table 1)
表1 粗山羊草 Aux/ IAA家族基因 Table 1 Aux/ IAA gene family in Ae. tauschii
基因 Gene
长度 Length (bp)
位点 Locus
基因组位置 Location
编码区序列 CDS
已定位序列 Mapped sequence
染色体 Chr.
AetIAA1
1012
scaffold63866
30422:31433
AEGTA00185
AT1D0116
1DS
AetIAA2
1886
scaffold7139
27496:29381
AEGTA27665
AT1D0209
1DS
AetIAA3
1367
scaffold41019
55515:56881
AEGTA14269
AT1D0278
1DS
AetIAA4
724
scaffold15242
4665:6198
AEGTA33259
AT1D0854
1DL
AetIAA5
421
scaffold47998
22106:22526
AEGTA14896
AT2D1713
2DL
AetIAA6
448
scaffold4386
204272:205064
AEGTA36582
AT2D2144
2DL
AetIAA7
1202
scaffold48637
5557:6758
AEGTA14144
AT3D2459
3DS
AetIAA8
4642
scaffold61519
16120:21472
AEGTA27928
AT3D2590
3DS
AetIAA9
4634
scaffold57491
7405:12539
AEGTA02358
AT3D3024
3DS
AetIAA10
1202
scaffold45297
19501:20702
AEGTA15903
AT3D3200
3DL
AetIAA11
1987
scaffold7072
197990:200210
AEGTA05621
AT4D3616
4DS
AetIAA12
4166
scaffold28516
31079:35244
AEGTA26128
AT5D4311
5DS
AetIAA13
831
scaffold36001
16911:17741
AEGTA22565
AT5D4872
5DL
AetIAA14
845
scaffold32507
25859:26703
AEGTA30995
AT5D4974
5DL
AetIAA15
2013
scaffold78860
21659:23671
AEGTA29089
AT5D4980
5DL
AetIAA16
774
scaffold11983
25370:26143
AEGTA30959
AT5D5091
5DL
AetIAA17
646
scaffold51919
31174:32211
AEGTA35747
AT5D5201
5DL
AetIAA18
1597
scaffold4121
160188:162577
AEGTA13779
AT6D5982
6DL
AetIAA19
986
scaffold96502
16230:17826
AEGTA32434
AT6D6008
6DS
AetIAA20
2193
scaffold96502
34944:35929
AEGTA09874
AT6D6008
6DS
AetIAA21
310
scaffold64080
56830:57139
AEGTA22007
AT7D6094
7DS
AetIAA22
2012
scaffold4040
9196:11207
AEGTA32199
AT7D6353
7DS
AetIAA23
551
scaffold58234
8830:9380
AEGTA29038
AT7D6529
7DS
AetIAA24
1442
scaffold55734
23879:25320
AEGTA02351
AT7D6725
7DL
AetIAA25
2249
scaffold129264
18851:21099
AEGTA05214
AT7D6800
7DL
AetIAA26
1526
scaffold21289
35533:37116
AEGTA04309
AT7D6841
7DL
AetIAA27
1015
scaffold354
44184:45385
AEGTA36436
AT7D7103
7DL
AetIAA28
2287
scaffold354
44222:46508
AEGTA22431
AT7D7103
7DL
表1 粗山羊草 Aux/ IAA家族基因 Table 1 Aux/ IAA gene family in Ae. tauschii
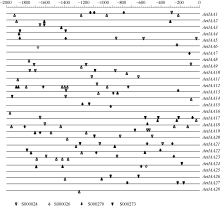
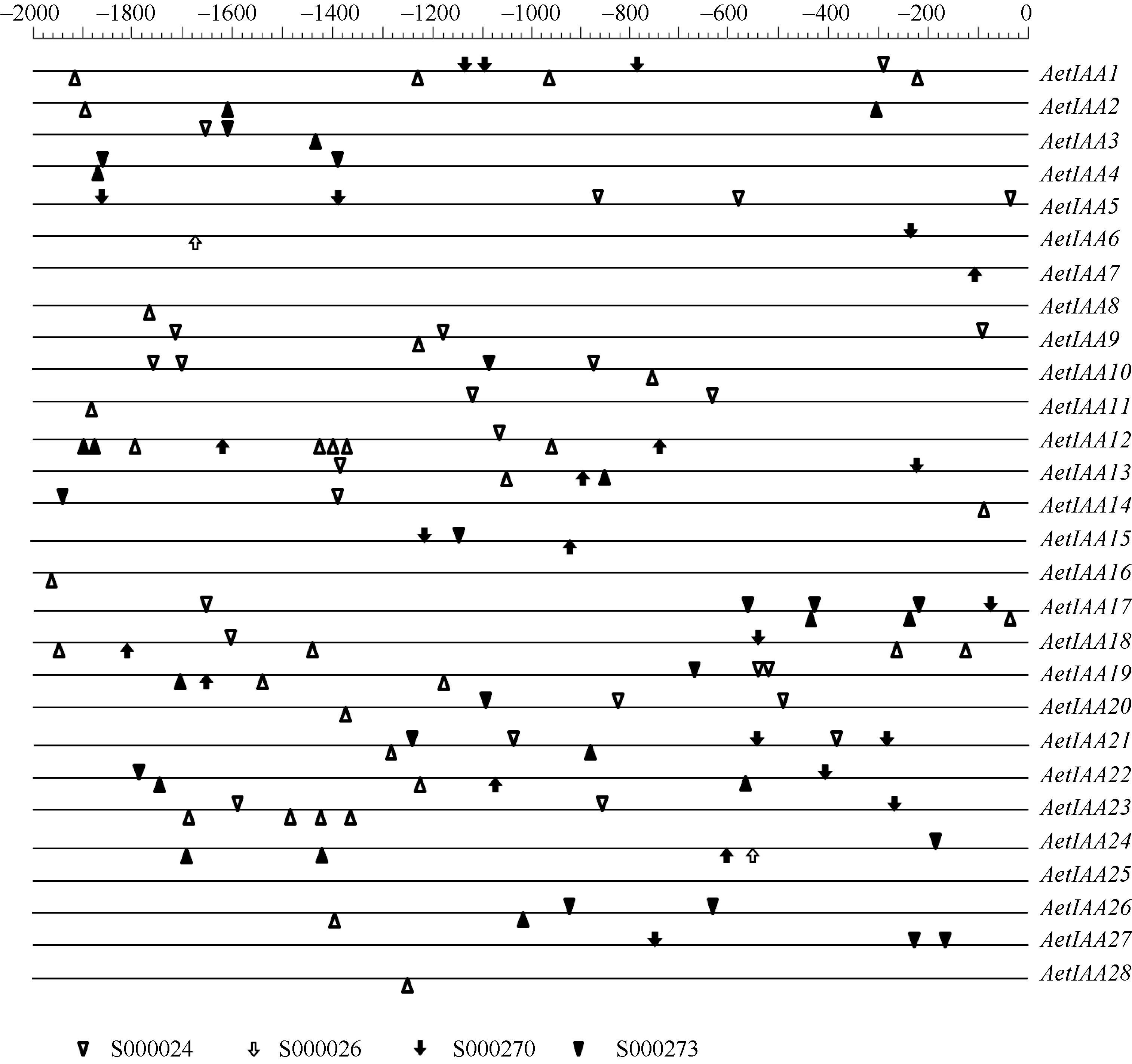
图5 粗山羊草 Aux/ IAA启动子区序列的顺式作用元件启动子序列用直线表示, 元件位于直线下方表示其调控方向相反。Fig. 5 cis-elements in the promoters of AetIAA genesThe sequences of promoters are indicated by line. Element located below the line means the regulation is reverse.
VannesteS, FrimlJ. Auxin: a trigger for change in plant deve-lopment. Cell, 2009, 136: 1005-1016[本文引用:1][JCR: 31.957]
[2]
WabnikK, Kleine-VehnJ, BallaJ, SauerM, NaramotoS, ReinöhlV, MerksR M, GovaertsW, FrimlJ. Emergence of tissue polarization from synergy of intracellular and extracellular auxin signaling. Mol Syst Biol, 2010, 21: 447[本文引用:1][JCR: 11.34]
[3]
LjungK. Auxin metabolism and homeostasis during plant deve-lopment. Development, 2013, 140: 943-950[本文引用:1][JCR: 6.208]
[4]
AbelS, TheologisA. Early genes and auxin action. Plant Physiol, 1996, 111: 9-17[本文引用:1][JCR: 6.555]
[5]
RoggL E, LasswellJ, BartelB. A gain-of-function mutation in IAA28 suppresses lateral root development. Plant Cell, 2001, 13: 465-480[本文引用:1][JCR: 9.251]
[6]
SongY, YouJ, XiongL. Characterization of OsIAA1 gene, a member of rice Aux/IAA family involved in auxin and brassinos-teroid hormone responses and plant morphogenesis. Plant Mol Biol, 2009, 70: 297-309[本文引用:1][JCR: 3.518]
[7]
KazanK, MannersJ M. Linking development to defense: auxin in plant-pathogen interactions. Trends Plant Sci, 2009, 14: 373-382[本文引用:1][JCR: 11.808]
[8]
StraderL C, ChenG L, BartelB. Ethylene directs auxin to control root cell expansion. Plant J, 2010, 64: 874-884[本文引用:1][JCR: 6.582]
[9]
KarenJ H, JaimeF M, Eve-Marie J. Integration of light and auxin signaling. Cold Spring Harb Perspect Biol, 2009, 1: 1-11[本文引用:1]
[10]
ReedJ W. Roles and activities of Aux/IAA proteins in Arabidopsis. Trends Plant Sci, 2001, 6: 420-425[本文引用:2][JCR: 11.808]
TiwariS B, WangX J, HagenG, GuilfoyleT J. Aux/IAA proteins are active repressors, and their stability and activity are modulated by auxin. Plant Cell, 2001, 13: 2809-2822[本文引用:1][JCR: 9.251]
[15]
KepinskiS, LeyserO. Auxin-induced SCF-TIR1-Aux/IAA interaction involves stable modification of the SCF/TIR1 complex. Proc Natl Acad Sci USA, 2004, 101: 12381-12386[本文引用:1][JCR: 9.737]
[16]
WalkerJ C, KeyJ L. Isolation of cloned cDNAs to auxin-respon-sive polyA RNAs of elongating soybean hypocotyl. Proc Natl Acad Sci USA, 1982, 79: 7185-7189[本文引用:2][JCR: 9.737]
[17]
PaulJ O, YokoO, José M A, April C, Chang C, Joseph R E, Beth H, Liu A, Courtney O, Hong Q, Alison S, Yu G X, Athanasios T. Functional genomic analysis of the AUXIN/INDOLE-3-ACETIC ACID gene family members in Arabidopsis thaliana. Plant Cell, 2005;17: 3282-3300[本文引用:2][JCR: 9.251]
[18]
JainM, KaurN, GargR, ThakurJ K, TyagiA K, KhuranaJ P. Structure and expression analysis of early auxin-responsive Aux/IAA gene family in rice (Oryza sativa). Funct Integr Geno-mics, 2006, 6: 47-59[本文引用:5]
[19]
UdayaC K, StephenP D, AmyM B, GeraldA T. Genome-wide analysis of Aux/IAA and ARF gene families in Populus trichocarpa. BMC Plant Biol, 2007, DOI: 10.1186/1471-2229-7-59[本文引用:2][JCR: 4.354]
[20]
WangY, DengD, BianY, Lü Y, Xie Q. Genome-wide analysis of primary auxin-responsive Aux/IAA gene family in maize (Zea mays L). Mol Biol Rep, 2010, 37: 3991-4001[本文引用:2][JCR: 2.506]
[21]
HanX, XuX, FangD D, ZhangT, GuoW. Cloning and expression analysis of novel Aux/IAA family genes in Gossypium hirsutum. Gene, 2012, 503: 83-91[本文引用:2][JCR: 2.196]
[22]
WuJ, PengZ, LiuS, HeY, ChengL, KongF, WangJ, LuG. Genome-wide analysis of Aux/IAA gene family in Solanaceae species using tomato as a model. Mol Genet Genomics, 2012, 287: 295-311[本文引用:3][JCR: 2.881]
[23]
GanD, ZhuangD, DingF, YuZ, ZhaoY. Identification and expression analysis of primary auxin-responsive Aux/IAA gene family in cucumber (Cucumis sativus). J Genet, 2013, 92: 513-521[本文引用:1][JCR: 0.876]
[24]
BhumicaS, ArchanaC, JitendraP K, ParamjitK. An early auxin-responsive Aux/IAA gene from wheat (Triticum aestivum) is induced by epibrassinolide and differentially regulated by light and calcium. J Exp Bot, 2006, 57: 4059-4070[本文引用:1][JCR: 5.242]
[25]
RachelB, ManuelS, MatthiasP, Gary L A B, Rosalinda D A, Alexand ra M A, Neil M, Melissa K, Arnaud K, Dan B, Suzanne K, Darren W, Martin T, Ian B, Gu Y, Huo N X, Luo M C, Sunish S, Bikram G, Sharyar K, Olin A, Paul K, Jan D, Richard M, Anthony H, Klaus F M, Keith J E, Michael W B, Hall N. Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature, 2012, 491: 705-710[本文引用:1][JCR: 38.597]
[26]
JiaJ Z, ZhaoS C, KongX Y, LiY R, ZhaoG Y, HeW M, AppelsR, PfeiferM, TaoY, ZhangX Y, JingR L, ZhangC, MaY Z, GaoL F, GaoC, SpannaglM, Mayer K F X, Li D, Pan S K, Zheng F Y, Hu Q, Xia X C, Li J W, Liang Q S, Chen J, Wicker T, Gou C Y, Kuang H H, He G Y, Luo Y D, Keller B, Xia Q J, Lu P, Wang J Y, Zou H F, Zhang R Z, Xu J Y, Gao J L, Middleton C, Quan Z W, Liu G M, Wang J, IWGSC, Yang H M, Liu X, He Z H, Mao L, Wang J. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature, 2013, 496: 91-95[本文引用:2][JCR: 38.597]
LuoM C, DealK R, AkhunovaE D, AkhunovaaA R, AndersonO D, AndersonJ A, BlakedN, CleggeM T, Coleman-Derrb D, Conley E J, Crossman C C, Dubcovskya J, Gill B S, Gu Y Q, Hadam J, Heod H Y, Huo N X, Lazo G, Ma Y, Matthewsg D E, McGuirea P E, Morrell P L, Qualseta C O, Renfrob J, Tabanao D, Talbertd L E, Tiana C, Tolenoe D M, Warburtonh M L, You F M, Zhang W, Dvoraka J. Genome comparisons reveal a dominant mechanism of chromosome number reduction in grasses and accelerated genome evolution in Triticeae. Proc Natl Acad Sci USA, 2009, 106: 15780-15785[本文引用:1][JCR: 9.737]
[32]
郭安源, 朱其慧, 陈新, 罗静初. GSDS: 基因结构显示系统. 遗传, 2007, 29: 1023-1026GuoA Y, ZhuQ H, ChenX, LuoJ C. GSDS: a gene structure display server. Hereditas (Beijing), 2007, 29: 1023-1026 (in Chinese with English abstract)[本文引用:1]
[33]
BaileyT L, BodenM, BuskeF A, FrithM, GrantC E, ClementiL, RenJ, LiW W, NobleW S. MEME SUITE: tools for motif discovery and searching. Nucl Acids Res, 2009, 37: 202-208[本文引用:1]
[34]
LarkinM A, BlackshieldsG, BrownN P, ChennaR, McGettigan, P A, McWilliam H, Valentin F, Wallace I M, Wilm A, Lopez R, Thompson J D, Gibson T J, Higgins D G. Clustal W and Clustal X version 2. 0. Bioinformatics, 2007, 23: 2947-2948[本文引用:1][JCR: 5.323]
KepinskiS, LeyserO. The Arabidopsis F-box protein TIR1 is an auxin receptor, Nature, 2005, 435: 446-451[本文引用:1][JCR: 38.597]
[37]
KimJ, HarterK, TheologisA. Protein-protein interactions among the Aux/IAA proteins. Proc Natl Acad Sci USA, 1997, 94: 11786-11791[本文引用:3][JCR: 9.737]
[38]
Colón-CarmonaA, ChenD L, YehK C, AbelS. Aux/IAA proteins are phosphorylated by phytochrome in vitro. Plant Physiol, 2000, 124: 1728-1738[本文引用:1][JCR: 6.555]
[39]
The International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature, 2009, 463: 763-768[本文引用:1][JCR: 38.597]
[40]
Martin-SanchezJ A, Gomez-ColmenarejoM, DelMoral J, SinE, MontesM J, González-BelinchónC, López-BrañaI, DelibesA. A new Hessian fly resistance gene (H30) transferred from the wild grass Aegilops triuncialis to hexaploid wheat. Theor Appl Genet, 2003, 106: 1248-1255[本文引用:1][JCR: 3.658]
[41]
NiJ, ZhuZ X, WangG H, ShenY X, ZhangY Y, WuP. Intragenic suppressor of Osiaa23 revealed a conserved tryptophan residue crucial for protein-protein interactions. PLoS One, 2014, 9: e85358[本文引用:1][JCR: 3.73]
McCartneyC A, SomersD J, HumphreysD G, LukowO, AmesN, NollJ, CloutierS, McCallumB D. Mapping quantitative trait loci controlling agronomic traits in the spring wheat cross RL4452 × ‘AC Domain’. Genome, 2005, 48: 870-883[本文引用:1][JCR: 1.668]
[44]
王瑞霞, 张秀英, 伍玲, 王瑞, 海林, 闫长生, 游光霞, 肖世和. 不同生态环境条件下小麦籽粒灌浆速率及千粒重QTL分析. 作物学报, 2008, 34: 1750-1756WangR X, ZhangX Y, WuL, WangR, HaiL, YanC S, YouG X, XiaoS H. QTL mapping for grain filling rate and thousand -grain weight in different ecological environments in wheat. Acta Agron Sin, 2008, 34: 1750-1756 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[45]
Audran-Delaland eC, BassaC, MilaI, RegadF, ZouineM, BouzayenM. Genome-wide identification, functional analysis and expression profiling of the Aux/IAA gene family in tomato. Plant Cell Physiol, 2012, 53: 659-672[本文引用:1][JCR: 4.134]
 , 郑军
, 郑军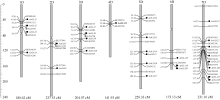







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}