关键词:叶片衰老; 叶绿体; 图位克隆; 糖基转移酶 Genetic Analysis and Fine Mapping of a Premature Leaf Senescence Mutant in Rice (Orzya sativaL.) ZHANG Tao1,2,**, SUN Yu-Ying3,**, ZHENG Jian-Min4,**, CHENG Zhi-Jun3, JIANG Kai-Feng1,2, YANG Li1,2, CAO Ying-Jiang1,2, YOU Shu-Mei1,2, WAN Jian-Min3, ZHENG Jia-Kui1,2,5,* 1 Rice and Sorghum Research Institute, Sichuan Academy of Agricultural Sciences / Key Laboratory of Southwest Rice Biology and Genetic Breeding, Ministry of Agriculture, Deyang 618000, China
2 Luzhou Branch of National Rice Improvement Center, Luzhou 646100, China
3 Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
4 Crop Research Institute, Sichuan Academy of Agricultural Sciences, Chengdu 610066, China
5 Bioengineering College, Chongqing University, Chongqing 400044, China
AbstractLeaf senescence induces degradation of chlorophyll and other macromolecules, reducing leaf photosynthetic capacity. This process is accompanied by the accumulation of reactive oxygen species (ROS), the decreasing of cell antioxidant enzyme (SOD, CAT, and APX) activity, and the increasing of aging related gene (SAG) expression, leading in early maturity and yield reduction. Therefore, studies on the genetic mechanism and gene function of premature senescence in rice, has the important effect and significance in genetic improvement of rice. PLS2 from space radiation mutation breeding project showed leaf senility, at booting stage. Compared with the wild type, in PLS2 the photosynthetic capacity decreased, the plant height, internode and panicle length shortened, tiller and effective tiller number reduced, number of grains per ear and seed setting rate were significantly lower, 1000-grain weight decreased, main panicle was stunted and grain-filling was not full. CAT activity decreased significantly in leaves, H2O2 accumulated, and the number of dead cell increased, chloroplast structures in leaves were worse, with more starches grains and osmiophilic granules. Dark treatment accelerated mutant leaf senescence, chloroplast ultrastructure was spheroidized. Using implicit localization population derived from PLS2/Shuhui 527 and PLS2/02428 thepls2 was located between markers RM14704 (8 674 283 bp) and SL-I-5 (8 758 394 bp) on chromosome 3, with physical distance of 84.11 kb, including 14 genes in the interval. Sequencing result showed that C was replaced with T in the position of 41 base pair of the ninth extron ofLOC_Os03g15840 leading to an exchange of Arg (R) is replaced by Cys(C).LOC_Os03g15840 coding a glycosyl transferase (GTs) may be the candidate gene ofpls2. These results provide a foundation for the further gene cloning and functional analysis ofpls2.
Keyword:Leaf senescence; Chloroplast; Map-based cloning; Glycosyltransferase1 Show Figures Show Figures
图1 野生型泸恢H103与突变体PLS2的表型A, B: 温室生长的六叶期野生型泸恢H103与突变体PLS2的表型; C: 野生型泸恢H103与突变体PLS2在抽穗期的表型; D: 野生型泸恢H103与突变体PLS2主穗的表型。Fig. 1 Phenotypes of wild type Luhui H103 and mutant PLS2 plantsA, B: Phenotypes of Luhui H103 and PLS2 plants at the 6th leaf stage; C: Phenotypes of WT and PLS2 plants at heading stage; D: Main panicle of wild type and PLS2 plants.
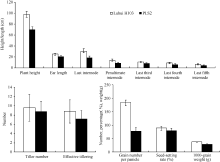
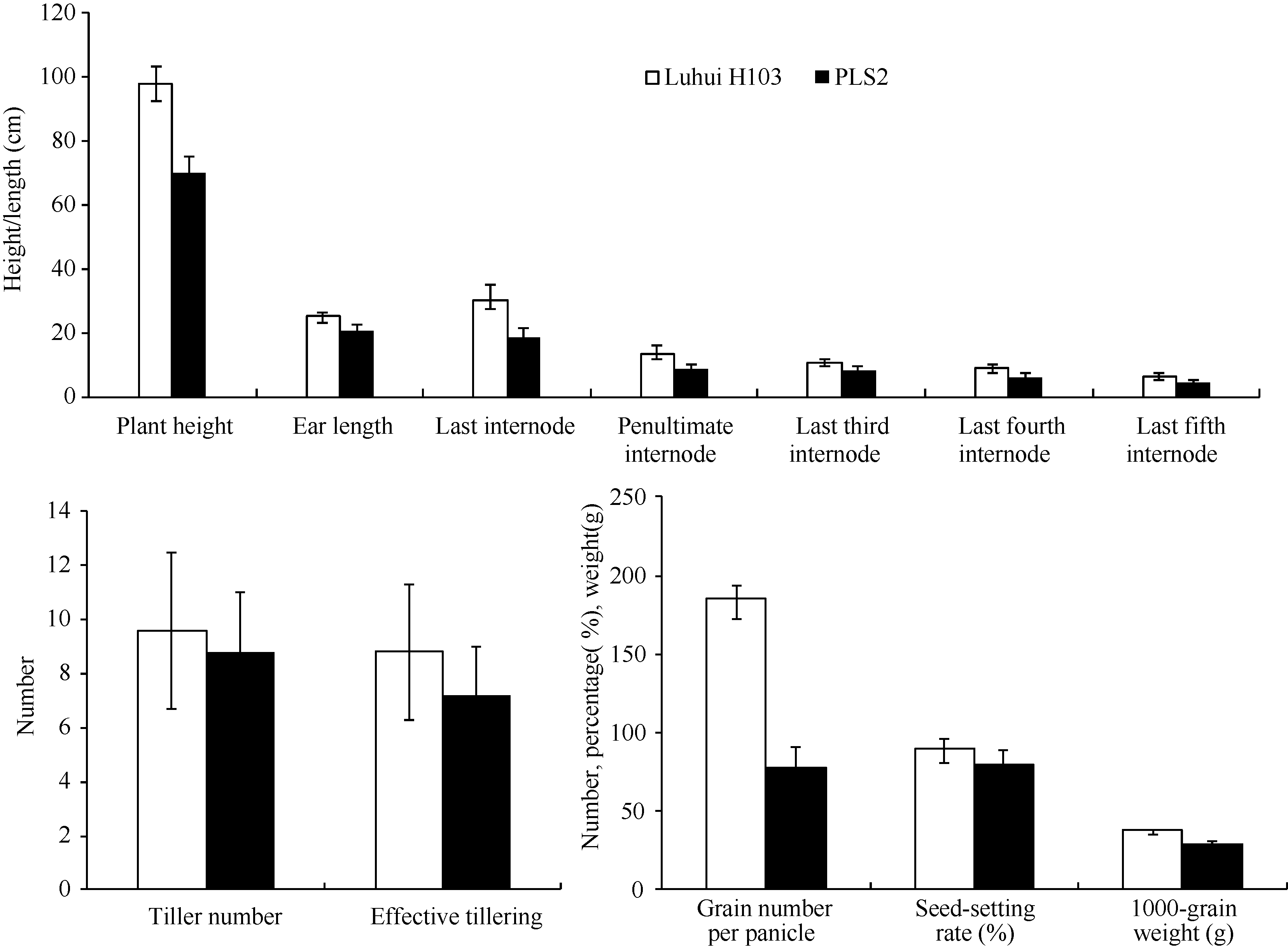
表1 野生型与突变体株高、穗长和节间的长度比较 Table 1 Comparison of plant height, ear length, and internodes between wild type and mutant (cm)
性状 Trait
地点 Site
泸恢H103 Luhui H103 (mean± SD)
PLS2 (mean± SD)
t值 t-value
株高 Plant height
北京 Beijing
97.6±5.7
70.2±5.0
2.6*
四川德阳 Deyang, Sichuan
117.4±4.4
93.4±3.9
2.9**
穗长 Ear length
北京 Beijing
25.0±1.3
20.5±2.2
0.3
四川德阳 Deyang, Sichuan
26.4±1.2
24.9±3.1
1.2
倒一节Last internodes
北京 Beijing
30.0±4.9
18.5±2.7
1.5
倒二节 Penultimate internodes
北京 Beijing
13.4±2.7
8.6±1.7
1.0
倒三节 Last third internodes
北京 Beijing
10.6±1.1
8.4±0.9
1.1
倒四节 Last fourth internodes
北京 Beijing
8.8±1.1
6.1±1.3
1.1
倒五节 Last fifth internodes
北京 Beijing
6.0±1.2
4.4±0.7
0.9
* t(α=0.05,8)=1.8595,** t(α=0.01,8)=2.8965.
表1 野生型与突变体株高、穗长和节间的长度比较 Table 1 Comparison of plant height, ear length, and internodes between wild type and mutant (cm)
表2 Table 2 表2(Table 2)
表2 野生型与突变体的单株产量、分蘖数、有效穗数、每穗粒数、结实率和千粒重比较 Table 2 Comparison of tiller number, effective tiller number, grain number per panicle, and seed-setting rate between wild type and mutant
性状 Trait
地点 Site
泸恢H103 Luhui H103 (mean± SD)
PLS2 (mean± SD)
t值 t-value
单株产量 Yield per plant (g)
四川德阳 Deyang, Sichuan
112.6±4.6
37.1±4.1
8.7**
分蘖数 Tiller number
北京 Beijing
9.6±2.9
8.8±2.2
0.2
有效穗 Effective tillering
北京 Beijing
8.8±2.5
7.2±1.8
0.4
四川德阳 Deyang, Sichuan
8.4±2.6
8.2±2.2
0
每穗粒数 Grain number per panicle
北京 Beijing
185.0±8.9
7.6±12.9
4.9**
四川德阳 Deyang, Sichuan
147.2±10.0
101.2±9.6
2.3*
结实率 Seed-setting rate (%)
北京 Beijing
89.7±6.0
79.4±9.0
0.7
四川德阳 Deyang, Sichuan
90.0±4.6
41.9±3.0
6.2**
千粒重 1000-grain weight (g)
北京 Beijing
37.5±0.2
28.4±3.0
2.2*
四川德阳 Deyang, Sichuan
35.4±0.4
33.6±0.4
2.3*
* t(α=0.05,8)=1.8595,** t(α=0.01,8)=2.8965.
表2 野生型与突变体的单株产量、分蘖数、有效穗数、每穗粒数、结实率和千粒重比较 Table 2 Comparison of tiller number, effective tiller number, grain number per panicle, and seed-setting rate between wild type and mutant
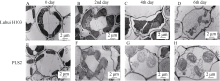
图3 生长4周的野生型泸恢H103与突变体PLS2在持续黑暗下的叶绿体降解情况上排为野生型叶绿体在黑暗处理前(A)、第2天(B)、第4天(C)和第6天(D)的降解情况, 下排为突变体叶绿体在黑暗处理前(E)、第2天(F)、第4天(G)和第6天(H)的降解情况。Fig. 3 Chloroplast degradation of wide type Luhui H103 and mutant PLS2 leaves in continuous dark treatmentA: WT before the treatment; B: WT in the second day of the treatment; C: WT in the fourth day of the treatment; D: WT in the sixth day of the treatment; E: PLS2 before the treatment; F: PLS2 in the second day of the treatment; G: PLS2 in the fourth day of the treatment; H: PLS2 in the sixth day of the treatment.
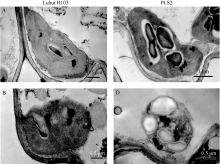
图4 野生型泸恢H103与突变体PLS2在抽穗之前的叶绿体结构左侧为野生型叶片的绿色部分(A)和黄色部分(B)叶绿体结构, 右侧为突变体叶片的绿色部分(C)和黄色部分(D)叶绿体结构。Fig. 4 Chloroplast structures of wide type Luhui H103 and mutant PLS2 leaves before heading stageA, B: Chloroplast structures of WT blade (corresponding leaf with C and D); C, D: Chloroplast structures of green and yellow parts in senescencing leaf of PLS2.
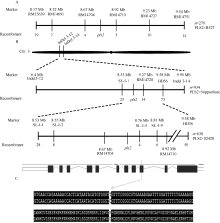
图5 pls2基因的图位克隆A, B: pls2基因的精细定位; C: 定位区间基因 LOC_Os03g15840的编码区存在碱基改变。Fig. 5 Map-based cloning of pls2 geneA, B: fine mapping of pls2 gene; C: sequence variations of genes in the mapping interval between WT and pls2.
图6 突变体基因 pls2在 LOC_Os03g15840的突变位点以及该基因的进化分析A: 基因 LOC_Os03g15840在不同水稻品种间的DNA序列比对分析; B: LOC_Os03g15840在不同物种间的氨基酸序列比对分析; C: pls2基因的进化分析。Fig. 6 Mutation position in pls2 of LOC_Os03g15840 compared with the homologous genes, and the phylogenetic tree of pls2A: Sequence BLAST of the LOC_Os03g15840 of pls2 between different rice varieties; B: Sequence BLAST of the LOC_Os03g15840 of pls2 between different plant species; C: Phylogenetic tree of pls2.
NavabpourS, MorrisK, AllenR, HarrisonE, A-H-Mackerness S, Buchanan-Wollaston V. Expression of senescence-enhanced genes in response to oxidative stress. J Exp Bot, 2003, 54: 2285-2292[本文引用:1][JCR: 5.242]
[2]
袁政, 张大兵. 植物叶片衰老的分子机制. 植物生理学通讯, 2002, 38: 417-422YuanZ, ZhangD B. The molecular mechanism of leaf senescence. Plant Physiol Commun, 2002, 38: 417-422 (in Chinese with English abstract)[本文引用:1][CJCR: 0.849]
EckardtN A. A new chlorophyll degradation pathway. Plant Cell, 2009, 21: 700-700[本文引用:1][JCR: 9.251]
[5]
LimP O, KimH J, NamH G. Leaf senescence. Annu Rev Plant Biol, 2007, 58: 115-136[本文引用:1][JCR: 25.962]
[6]
ChaK W, LeeY J, KohH J, LeeB M, NamY W, PaekN C. Isolation, characterization, and mapping of stay green mutant in rice. Theor Appl Genet, 2002, 104: 526-532[本文引用:1][JCR: 3.658]
[7]
LeeR H, LinM C, ChenS C. A novel alkaline α-galactosidase gene is involved in rice leaf senescence. Plant Mol Biol, 2004, 55: 281-295[本文引用:1][JCR: 3.518]
[8]
AnsariM I, LeeR H, ChenS C. A novel senescence-associated gene encoding GABA: pyruvate transaminase is up-regulated during rice leaf senescence. Physiol Plant, 2005, 123: 1-8[本文引用:1][JCR: 6.555]
[9]
KongZ S, LiM N, YangW Q, XuW Y, XueY B. A novel nuclear localized CCCH-type zinc finger protein, OsDOS, is involved in delaying leaf senescence in rice. Plant Physiol, 2006, 141: 1376-1388[本文引用:1][JCR: 6.555]
[10]
JiangH W, LiM R, LiangN T, YanH B, WeiY B, XuX L, LiuJ, XuZ F, ChenF, WuG J. Molecular cloning and function analysis of the stay green gene in rice. Plant J, 2007, 52: 197-209[本文引用:1][JCR: 6.582]
[11]
ParkS Y, YuJ W, ParkJ S, LiJ J, YooS C, LeeN Y, LeeS K, JeongS W, SeoH S, KohH J, JeonJ S, ParkY, PaekN C. The senescence-induced stay green protein regulates chlorophyll degradation. Plant Cell, 2007, 19: 1649-1664[本文引用:1][JCR: 9.251]
[12]
KusabaM, ItoH, MoritaR, IidaS, SatoY, FujimotoM, KawasakiS, TanakaR, HirochikaH, NishimuraM, TanakaA. Rice NON-YELLOW COLORINGI is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell, 2007, 19: 1362-1375[本文引用:1][JCR: 9.251]
[13]
WuZ M, ZhangX, HeB, DiaoL P, ShengS L, WangJ L, GuoX P, SuN, WangL F, JiangL, WangC M, ZhaiH Q, WanJ M. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:1][JCR: 6.555]
[14]
QiaoY L, JiangW Z, LeeJ H, ParkB S, ChoiM S, PiaoR H, WooM O, RohJ H, HanL Z, PaekN C, SeoH S, KohH J. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit 1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol, 2009, 185: 258-274[本文引用:1][JCR: 6.736]
[15]
LiF Z, HuG C, FuY P, SiH M, BaiX M, SunZ X. Genetic analysis and high-resolution mapping of premature senescence gene Pse(t) in rice (Oryza sativa L. ). Genome, 2005, 48: 738-746[本文引用:1][JCR: 1.668]
[16]
WangJ, WuS J, ZhouY, ZhouL H, XuJ F, HuJ, FangY X, GuM H, LiangG H. Genetic analysis and molecular mapping of a presenescing leaf gene pls1 in rice (Oryza sativa L. ). Chin Sci Bull, 2006, 51: 2986-2992[本文引用:1][CJCR: 0.95]
[17]
YanW Y, YeS, JinQ S, ZengL J, PengY, YanD W, YangW B, YangD L, HeZ H, DongY J, ZhangX M. Characterization and mapping of a novel mutant sms1 (senescence and male sterility 1) in rice. J Genet Genomics, 2010, 37: 47-55[本文引用:1][JCR: 2.076][CJCR: 1.323]
[18]
FangL K, LiY F, GongX P, SangX C, LingY H, WangX W, CongY F, HeG H. Genetic analysis and high-resolution mapping of pls3 (presenescing leaf 3) in rice. Chin Sci Bull, 2010, 55: 1676-1681[本文引用:1][CJCR: 0.95]
[19]
YangY L, RaoY C, LiuH J, FangY X, DongG J, HuangL C, LengY J, GuoL B, ZhangG H, HuJ, GaoZ Y, QianQ, ZengD L. Characterization and fine mapping of an early senescence mutant (es-t) in Oryza sativa L. Chin Sci Bull, 2011, 56: 2437−2443[本文引用:1][CJCR: 0.95]
[20]
杜青, 方立魁, 桑贤春, 凌英华, 李云峰, 杨正林, 何光华, 赵芳明. 水稻叶尖早衰突变体lad的形态、生理分析与基因定位. 作物学报, 2012, 38: 168-173DuQ, FangL K, SangX C, LingY H, LiY F, YangZ L, HeG H, ZhaoF M. Analysis of phenotype and physiology of leaf apex dead mutant (lad) in rice and mapping of mutant gene. Acta Agron Sin, 2012, 38: 168-173 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[21]
徐芳芳, 桑贤春, 任德勇, 唐彦强, 胡宏伟, 杨正林, 赵芳明, 何光华. 水稻早衰突变体esl2的遗传分析和基因定位. 作物学报, 2012, 38: 1347-1353XuF F, SangX C, RenD Y, TangY Q, HuH W, YangZ L, ZhaoF M, HeG H. Genetic analysis and gene mapping of early senescence leaf mutant esl2 in rice. Acta Agron Sin, 2012, 38: 1347-1353 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[22]
苗润隆, 蒋钰东, 廖红香, 徐芳芳, 何光华, 杨正林, 赵芳明, 桑贤春. 水稻早衰突变体esl3的鉴定与基因定位. 作物学报, 2013, 39: 862-867MiaoR L, JiangY D, LiaoH X, XuF F, HeG h, YangZ L, ZhaoF M, SangX C. Identification and gene mapping of rice early senescent leaf (esl3) mutant. Acta Agron Sin, 2013, 39: 862-867 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[23]
王复标, 黄福灯, 程方民, 李兆伟, 胡东维, 潘刚, 毛愉婵. 水稻生育后期叶片早衰突变体的光合特性与叶绿体超微结构观察. 作物学报, 2012, 38: 871-879WangF B, HuangF D, ChengF M, LiZ W, HuD W, PanG, MaoY C. Photosynthesis and chloroplast ultra-structure characteristics of flag leaves for a premature senescence rice mutant. Acta Agron Sin, 2012, 38: 871-879[本文引用:1][CJCR: 1.667]
[24]
UndanJ R, TamiruM, AbeA, YoshidaK, KosugiS, TakagiH, YoshidaK, KanzakiH, SaitohH, FekihR. Mutation in OsLMS, a gene encoding a protein with two double-strand ed RNA binding motifs, causes lesion mimic phenotype and early senescence in rice (Oryza sativa L. ). Genes Genet Syst, 2012, 87: 169-179[本文引用:1][JCR: 1.132]
[25]
LinA H, WangY Q, TangJ Y, XueP, LiC L, LiuL C, HuB, YangF Q, LoakeG J, ChuC C. Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide induced leaf cell death in rice. Plant Physiol, 2012, 158: 451−464[本文引用:1][JCR: 6.555]
[26]
张涛, 郑家奎, 蒋开锋, 郑建敏, 杨乾华, 杨莉, 万先齐, 曹应江. 水稻航天衰老突变体基因psl2的表型和遗传分析. 分子植物育种, 2010, 8: 245-251ZhangT, ZhengJ K, JiangK F, ZhengJ M, YangQ H, YangL, WanX Q, CaoY J. Phenotypes and genetic analysis of a senescence mutant by aeronautics in rice. Mol Plant Breed, 2010, 8: 245-251(in Chinese with English abstract)[本文引用:2][CJCR: 1.23]
[27]
张涛. 水稻糙米蛋白质含量的QTL定位及香稻的资源研究. 四川农业大学博士学位论文, 四川雅安, 2007. p25ZhangT. QTL Mapping of Brown Rice Protein Content in a RIL Population of Rice and Analysis of Genetic Diversity of Aromatic Rice Varieties. PhD Dissertation of Sichuan Agricultural University, Sichuan, China, 2007. p25 (in Chinese with English abstract)[本文引用:1]
[28]
RogersS O, BendichA J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol Biol, 1985, 5: 69-76[本文引用:1][JCR: 3.518]
[29]
LinQ B, WangD, DongH, GuS H, ChengZ J, GongJ, QinR Z, JiangL, LiG, WangJ L, WuF Q, GuoX P, ZhangX, LeiC L, WangH Y, WanJ M. Rice APC/CTE controls tillering by mediating the degradation of MONOCULM1. Nat Commun, 2012, 3: 752, DOI: 10.1038/ncomms1716www.nature.com/natureco-mmu-nications[本文引用:1][JCR: 10.015]
[30]
Buchanan-WollastonV, PageT, HarrisonE, BreezeE, LimP O, NamH G, LinJ F, WuS H, SwidzinskiJ, IshizakiK, LeaverC J. Comparative transcriptome analysis reveals significant differences in gene expression and signaling pathways between deve-lopmental and dark/starvation induced senescence in Arabidopsis. Plant J, 2005, 42: 567-585[本文引用:1][JCR: 6.582]
[31]
马跃芳, 陆定志. 灌水方式对杂交水稻衰老及生育后期一些生理活性的影响. 中国水稻科学, 1990, 4: 56-62MaY F, LuD Z. Effect of irrigation modes on the senescence and physiological activity in hybrid rice after heeding. Chin J Rice Sci, 1990, 4: 56-62 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[32]
WooH H, JeongB R, KooK B, ChoiJ W, HirschA M, HawesM C. Modifying expression of closely related UDP-glycosyltran-s-ferases from pea and Arabidopsis results in altered root deve-lopment and function. Physiol Plant, 2007, 130: 250-260[本文引用:1][JCR: 6.555]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}