 , 袁守江
, 袁守江*通讯作者(Corresponding authors): 李学勇, E-mail:lixueyong@caas.cn, Tel: 010-82107409; 袁守江, E-mail:ysj868@sina.com, Tel: 15615973691 收稿日期:2014-07-06 基金:
摘要
关键词:水稻; 类树状突变体; 出叶间隔期; 基因定位;
Genetic Analysis and Gene Fine Mapping of Rice Leafy Head Mutant
WANG Hai-Feng
, YUAN Shou-JiangFund:
Abstract
Keyword:Rice; Leafy head mutant; Plastochron; Gene mapping;
Show Figures
Show Figures



叶片是植物光合作用的主要器官, 也是植物株型的重要组成部分。叶片的发育起始于茎顶端分生组织周边区的起始细胞, 这些细胞的不断分化形成叶原基, 之后发育成完整的叶片[ 1]。这是一个复杂的生物学过程, 除内在遗传因素占主导作用外, 外界环境条件也会影响叶片的发育[ 2]。近年来, 随着分子生物学和突变体研究技术的发展, 借助5种模式植物即双子叶植物拟南芥( Arabidopsis thaliana)、烟草( Nicotiana tabacum)、金鱼草( Antirrhinum majus)、番茄( Lycopersicon esculentum)和单子叶植物玉米( Zea mays), 植物叶片发育的相关分子机制和调控网络研究已取得重大进展, 发现了多种与叶片发育相关的基因、miRNA和植物激素[ 3, 4]。
叶序和出叶间隔期是叶片生长发育的基本方面和物种特性。通过对叶序或出叶间隔期突变体的研究, 可以帮助我们了解叶片的形成机制。水稻的叶序和出叶间隔期也是其农艺性状的重要方面。目前已经从不同植物中克隆了控制出叶间隔期的几个基因, 如水稻 PLASTOCHRON1( PLA1)[ 5]、 PLASTOC-HRON2( PLA2) /LEAFY HEAD2( LHD2)[ 6, 7]和 PLAST-OCHRON3( PLA3)/ GOLIATH( GO)[ 8], 玉米 terminal ear1( te1)[ 9]以及拟南芥 ALTERED MERISTEM PROGRAM1( AMP1)、 PHYTOCHROME B ( PHYB)和 SERRATE( SE)[ 10, 11, 12]。功能丧失突变体 pla1、 pla2/lhd2、 pla3、 te1和 amp1均表现出出叶间隔期缩短的现象, 而 phyB和 se的出叶间隔期却长于野生型。 pla1的出叶时间间隔较野生型缩短了一半, 株高和叶片大小也减小到野生型的一半, 但进入生殖生长的时间并未发生明显变化[ 5]。 pla2/lhd2表现为出叶间隔变短, 植株矮化, 分蘖数减少且不能正常进行营养生长向生殖生长的转变[ 6, 7]。 pla3出叶间隔缩短, 不能正常进行营养生长向生殖生长的转变, 种胚增大、种子萌发后顶端分生组织缺陷和叶形态异常。
我们在日本晴的EMS (ethylmethane sulfonate)诱变群体中发现了一个水稻类树状突变体, 该突变体在幼苗期出叶速度快, 叶片变窄、变短, 叶片与叶鞘的角度比野生型大, 植株矮化、分蘖数减少、不能正常进行生殖生长。遗传分析和基因定位结果表明, F2群体中正常株和突变株符合3∶1的分离比例, 说明该性状受1对隐性基因控制。将此基因精细定位于88 kb的物理距离范围内, 这个区间包含11个基因, 在水稻中已报道的与本突变体相关的基因为 LHD2/PLA2。因此, 参考 PLA2的基因设计引物对本突变体进行基因测序。测序结果表明, 该突变体为 PLA2第4内含子的第5位碱基由G突变成A的单碱基突变, 为 pla2/lhd2的等位突变体。
1 材料与方法1.1 试验材料通过甲基磺酸乙酯(EMS)诱变粳稻品种日本晴获得一个稳定遗传的类树状突变体 s2-21, 该突变表现为出叶间隔变短、矮化、分蘖数减少、不能正常进行生殖生长。
1.2 F2分离群体的构建与遗传分析 以突变体 s2-21的杂合个体作为母本, 与Dular杂交获得 s2-21/Dular的F1代。2010年在海南三亚单株种植F1, 分单株收获F2代种子。2011年在山东水稻研究所济宁试验场单株种植F2。从 s2-21/Dular的F2分离群体中选取突变体植株用于初步定位和精细定位, 同时统计F2群体中的正常和突变个体的株数用于遗传学分析。
1.3 DNA提取和基因初步定位从 s2-21突变体及父母亲本每株取嫩叶0.1 g, 采用CTAB (十六烷基三甲基溴化铵)法[ 13]提取总DNA, 用于PCR分析。
以BSA法[ 14](bulked segregant analysis)构建F2代正常个体和突变个体的混池(20株DNA等量混合), 利用InDel标记对分离群体的2个亲本进行多态性分析, 并进一步利用多态性标记分析2个混池间的多态性, 找出可能与突变体基因连锁的标记, 然后用这些连锁标记分析F2群体中各突变单株。
所用的标记为本实验室根据籼、粳稻之间插入或缺失差异设计的, 共170个。参照Panaud等[ 15]的PCR体系, 反应产物在220 V电压下以8%聚丙烯酰胺凝胶电泳1 h, 再经硝酸银染色后照相分析。
1.4 发展新标记与精细定位根据初步定位结果, 利用本实验室已测序的籼稻品种Dular全基因组序列(待发表)与NCBI (http://www.ncbi.nlm.nih.gov/)上提供的粳稻品种日本晴基因组序列比对, 选取在初步定位区间内插入或缺失10~30 bp的多态性区域, 设计扩增产物为200 bp左右的InDel标记, 合成的引物先对两亲本进行多态性筛选。用扩增良好并表现多态性的引物分别对定位群体中正常植株和突变植株进行分析, 用于精细定位。
1.5 连锁分析根据电泳结果, 含母本日本晴带型的单株标记为0, 含父本Dular带型的单株标记为2, 含父母本带型的标记为1。利用MapMaker3.0作图构建目标区域的分子标记连锁图并计算遗传距离。
1.6 基因测序根据突变基因精细定位的区段结果, 通过Rice Genome Annotation Project (水稻基因组注释系统)网站(http://rice.plantbiology.msu.edu/)进行预测, 查看此区段内的开放阅读框, 初步了解每个基因的功能, 并对候选基因进行测序。
2 结果与分析2.1 突变体的表型分析与野生型日本晴相比, 突变体 s2-21出叶速度快, 总叶片数增多, 叶面积减小, 节间缩短, 分蘖数减少, 植株变矮, 而且不能正常进行生殖生长(图1)。
从图2可以看出, s2-21苗期的出叶速度是野生型的2倍, 且在野生型的抽穗期时, 突变体 s2-21总叶片数是野生型的2倍。
2.2 遗传分析突变体 s2-21与Dular杂交, F1个体表现正常, F2群体中正常表型和突变体表型的个体分别为354株和103株, 其分离比符合3∶1 (χ2=1.414 < χ20.05=3.84, 0.05< P<0.10), 表明该突变体的性状受1对隐性基因控制。
图1
Fig. 1
| Figure OptionViewDownloadNew Window | |
 | 图1 野生型日本晴和突变体 s2-21的表型A: 14 d秧龄植株表型; B、C: 抽穗期表型; D: 抽穗期节间表型; 左侧为日本晴, 右侧为 s2-21突变体; E: 抽穗期突变体 S2-21表型。A标尺为1 cm; B、C、D标尺为5 cm; E标尺为2 cm。Fig. 1 Phenotypes of wild type Nipponbare and s2-21A: phenotype of the 14 d old seedling; B and C: phenotype at the heading stage; D: stems of plants at the heading stage. Left: Nipponbare; Right: s2-21mutant .E: phenotype of the s2-21 mutant at the heading stage. Bar = 1 cm (in A); bar = 5 cm (in B, C, D); bar = 2 cm (in E). |
图2
Fig. 2
| Figure OptionViewDownloadNew Window | |
 | 图2 野生型和 s2-21农艺性状比较A: 苗期叶片生长动态; B: 成熟期主茎叶片数和植株分蘖数; C: 苗期和抽穗期株高。B和C中数值为平均值±标准误差, n= 10。Fig. 2 Agronomic traits of wild type and s2-21A: comparison of leaf development between the wild-type and the s2-21 mutant at the seedling stage. B:the number of main stem leaves and the number of tillers at the maturation stage. C: plant height at the seedling stage and the heading stage (value: mean ± SE, n = 10). |
2.3 基因的初步定位利用水稻12条染色体上的170个InDel标记和已建成的突变体混池进行连锁筛选表明, 该突变体基因与第一染色体上的C1-15和S1-17连锁。进一步利用这2个标记对45个F2突变体单株初步验证发现, 与这2个标记的遗传距离分别为26.0 cM和1.9 cM, 突变基因被初步定位在水稻第1染色体上的C1-15和S1-17之间(图3)。
2.4 s2-21突变基因的精细定位和候选基因测序 为进一步精细定位突变基因, 将F2群体扩大到737株突变个体, 使用两侧的标记C1-15和S1-17-1进行初筛, 分别有206个和29个交换单株。根据粳稻品种日本晴和籼稻品种Dular基因组序列之间的差异, 又发展了29个标记, 其中有6个在两亲本间具有多态性, 即w5、w10、w23、w24、w25和w26 (表1), 在这6个标记处的交换单株个数分别为151、88、21、4、1和1, 最终将 s2-21突变体基因定位在W25和W26之间, 这2个标记分别在BAC克隆AP006531和AP003416上, 物理距离约为88 kb (图3)。利用Rice Genome Annotation (水稻基因组注释系统)网站(http://rice.plantbiology.msu.edu/)预测此区域有11个基因, 其中编号为LOC_Os01g68000的基因突变后产生的表型与 s2-21相似, 并被命名为 LHD2/PLA2[ 6, 7]。因此, 参考 PLA2基因设计引物对本突变体进行基因测序表明, 突变体 s2-21中的 PLA2基因第4内含子的第5位碱基由G突变成A。
图3
Fig. 3
| Figure OptionViewDownloadNew Window | |
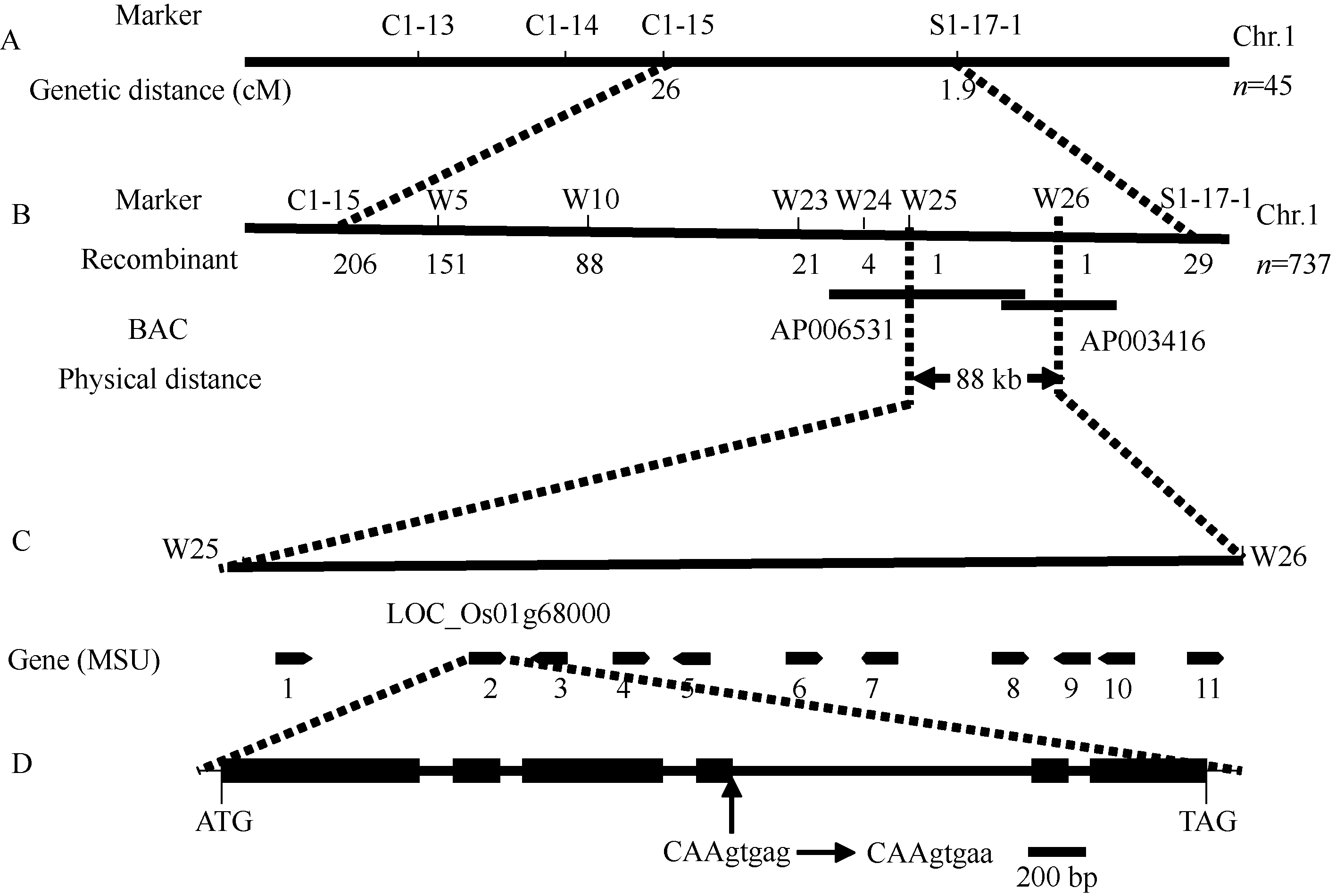 | 图3 s2-21在水稻第1染色体上的精细定位A: 基因的初步定位; B: 基因的精细定位(数值表示发生的交换单株个数); C: 定位区间的预测基因和突变基因; D: PLA2基因结构, 该基因包含6个外显子和5个内含子, 箭头指示突变体的突变位点。Fig. 3 Fine mapping of s2-21 on chromosome 1A: primary mapping of s2-21mutant; B: fine mapping of mutant allele region (the numbers of recombination events were shown); C: the candidate region of the s2-21gene and the predicted OFRs in this interval; D: struture of the s2-21gene. It consists of six exons (filled rectangles) and five introns (black line). Arrows indicate the mutation site in the s2-21 mutant. |
表1
Table 1
表1(Table 1)
| 表1 用于基因定位的分子标记 Table 1 Molecular markers used for gene mapping |
3 讨论用自然突变、理化诱变、T-DNA插入诱变及转座子插入诱变等方法已创制了10多万份水稻突变体, 极大地富了水稻功能基因组学的研究对象[ 16]。本研究通过EMS诱变处理粳稻品种日本晴获得了1个稳定遗传的类树状突变体 s2-21, 并对其进行了遗传分析和精细定位。该突变体与野生型日本晴相比, 出叶速度变快, 总叶片数增多, 叶面积减小, 节间缩短, 分蘖数减少, 植株变矮, 而且不能正常进行生殖生长。
叶片发育包括叶形、叶面积和叶序, 是影响植物株型的决定性因素。因此水稻叶片的起始发育模式对其株型尤为重要。叶片的发育模式取决于2个相邻叶原基形成的间隔期和叶序, 所以这两个方面异常的突变体不仅有助于研究叶片形成的机理, 而且可以丰富株型改良的手段, 进而提高水稻产量。Kauzumaru等[ 5]和Taiji等[ 6]分别从水稻突变体中发现 PLA1和 PLA2是调控水稻叶序发育的重要因素, 这2个基因均能缩短第1个与第2个叶原基形成之间的间隔期。熊国胜等[ 7]从云稻32和TN1的杂交后代中筛选出自然突变体 lhd2-1和 lhd2-3, 并且用EMS诱变的方法从日本晴中筛选出了突变体 lhd2-2, 并将 LHD2定位于第1染色体长臂。与野生型比较, 这些突变体在苗期就表现出更快的出叶速率, 叶面积和植株生长量也减小。
水稻生育期决定品种的地区和季节适应性, 这一直都是水稻育种研究的目标之一。水稻生育期分为营养生长和生殖生长2个时期, 不同水稻品种之间营养生长期的时间相差很大。所以水稻品种生育期的不同主要是营养生长期变化所致。目前已鉴定了一些控制水稻生育期的基因。经典遗传研究表明, 营养生长期主要由2个或3个Ef基因控制, 感光性由 Se1、 Se2、 Se3、 Se4、 Se5、 Se6、 Se7及 E1、 E2、 E3等一系列的基因控制。 E1和 Se1两个主效感光性基因是影响水稻抽穗期的主要因子, 其他一些基因已用形态标记及同工酶标记进行了定位。近10年来, 约536个水稻抽穗期QTL被定位, 其中 Hd1、 Hd6和 Ehd1等多个数量性状基因已被克隆[ 17]。 PLA1和 PLA2/ LHD2这两个控制叶片发育的基因均能延长水稻的营养生长期, 导致水稻不能正常向生殖生长期转变[ 5, 6, 7]。 lhd3突变体在短日照条件下, 在营养生长后期上部节间会继续长出叶片和高位分蘖而基部分蘖不受影响, 导致营养期延长, 抽穗期延迟; 但是最终不会影响抽穗结实。本文报道的突变体只进行营养生长而不能进行生殖生长。通过精细定位及测序发现该突变基因为 PLA2的等位基因, 是 PLA2基因的第4个内含子的第5位碱基由G突变为A。在植物研究中, 由于内含子突变而导致表型变化的现象还未见报道。我们推测该内含子突变可能影响mRNA前体剪接的准确性或效率, 从而影响 PLA2基因的准确表达。
4 结论以EMS化学诱变粳稻品种日本晴, 获得了1个类树状突变体 s2-21, 其出叶间隔期变短、植株矮化、不能正常进行生殖生长。该性状由1对隐性基因控制, 被定位在第1染色体长臂上88 kb的区间内, 测序发现 PLA2基因的第4个内含子的第5位碱基由G突变为A。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] |

{kind=link}
{kind=link}
{kind=link}