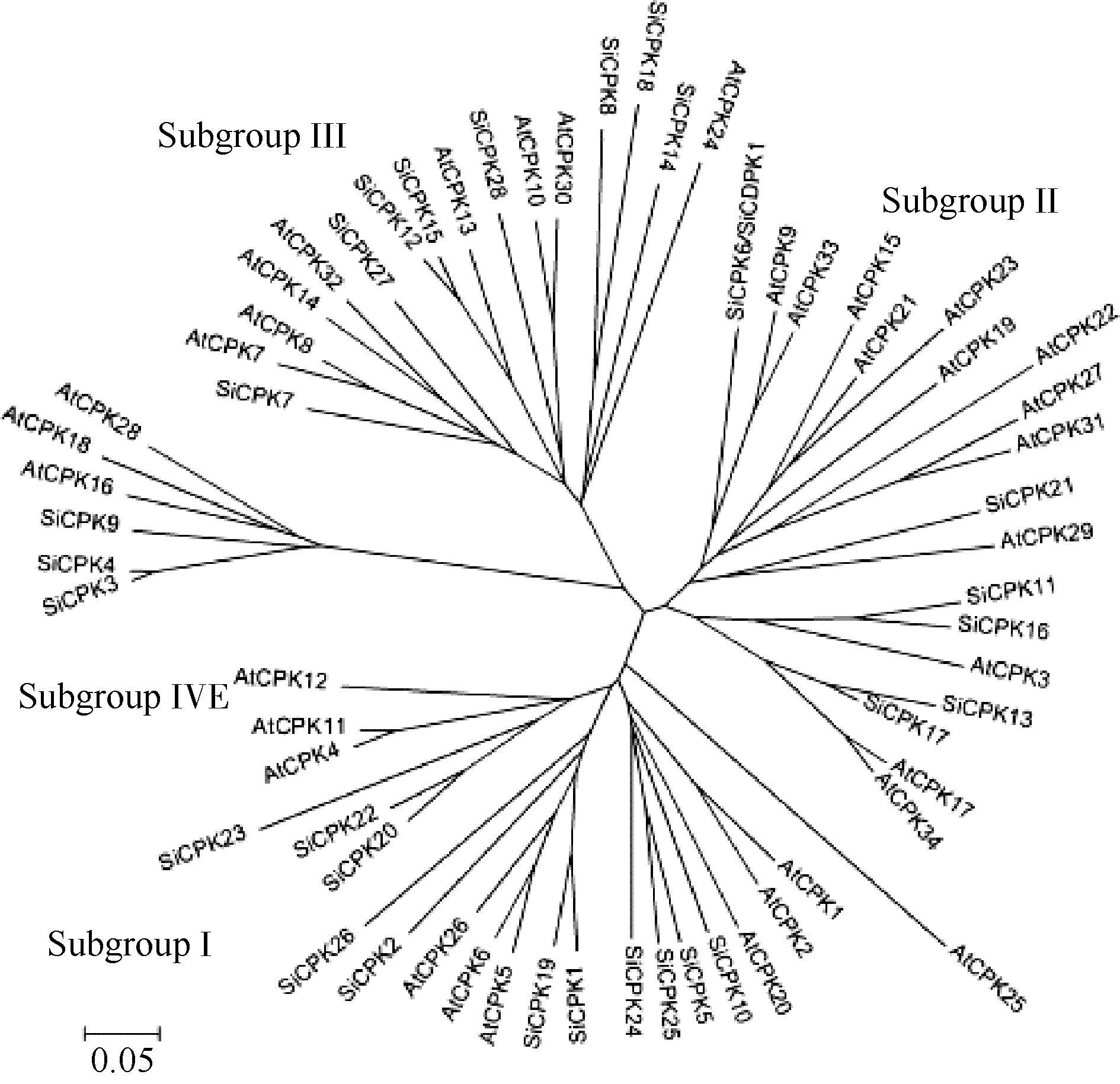
关键词:谷子; CDPK; 逆境胁迫; 钙信号 Cloning and Expression Analysis of a Calcium-Dependent Protein Kinase GeneSiCDPK1 inSetaria italica YU Qin-Yang, YIN Heng, AN Li-Jia, LI Wen-Li* School of Life Science and Biotechnology, Dalian University of Technology, Dalian 116023, China Fund: AbstractCDPK (calcium-dependent protein kinase) is a kind of essential calcium sensors and calcium responders, which plays important roles in response to various abiotic stresses in plants. In this research, we cloned the abiotic stress response relatedCDPK gene with RT-PCR inSetaria italica, designated asSiCDPK1 (GenBank accession number KC249975.1). We predicted 28CDPKs in theSetaria italicagenome by using the knownArabidopsis CDPK sequences as query sequences. The phylogenetic analysis showed theSiCDPK gene family was divided into four subgroups, in whichSiCDPK1belongs to subgroup II. The ORF ofSiCDPK1contains 1596 bp, which encodes 531 amino acids. The predicted protein molecular weight is 59.5 kD and pI is 5.94. SiCDPK1 has conserved protein domains of CDPK. The regulatory element analysis of promoter inSiCDPK1 showed a lot ofcis-acting elements associated with different abiotic stresses. The RT-PCR results showed thatSiCDPK1 was induced by PEG, ABA, salinity and drought. The research onSiCDPK1paves a way for unraveling the mechanism of abiotic stresses in plants.
图1 SiCDPK1 基因PCR扩增条带1~4: SiCDPK1在53.0、54.8、56.6和58.4℃下PCR结果; M: DL2000(TaKaRa)。1~4: SiCDPK1在53.0、54.8、56.6和58.4℃下PCR结果; M: DL2000(TaKaRa)。Fig. 1 PCR results of SiCDPK1 gene1-4: PCR products of SiCDPK1 gene at the temperature of 53.0, 54.8, 56.6, and 58.4℃ respectively; M: DL2000 (TaKaRa).
图3 SiCDPK1基因序列及推导氨基酸序列分析阴影部分为蛋白激酶区, 加粗部分为钙离子结合区, 两者中间为自抑连接区, 其中虚线框内为自抑片段, 剩余部分为可变区。下画线显示的是EF hand结构域。序列开头框内为豆蔻酰化保守序列MGXXXSXX, 其中G为豆蔻酰化位点, C为棕榈酰化位点。Fig. 3 Analysis of SiCDPK1 gene sequence and deduced amino acid sequenceThe shadow shows the protein kinase domain, and the bold shows the calmodulin-like domain. The junction domain is between the protein kinase domain and the calmodulin-like domain, in which the autoinhibitory domain is shown by dashed box. The rest is variable domain. EF hand is underlined. The myristoylation conserved domain is MGXXXSXX, which is shown by the box at the beginning of the text. G is myristoylation site and C is palmitoylation site.
图5 SiCDPK1启动子顺式调控元件示意图直线上方为(+)链调控元件, 下方为(-)链调控元件。Fig. 5 Schematic diagram of cis-acting element in the promoter region of SiCDPK1The element above the line is in (+)-sense strand, and the element below the line is in (-)-sense strand.
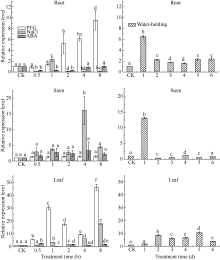
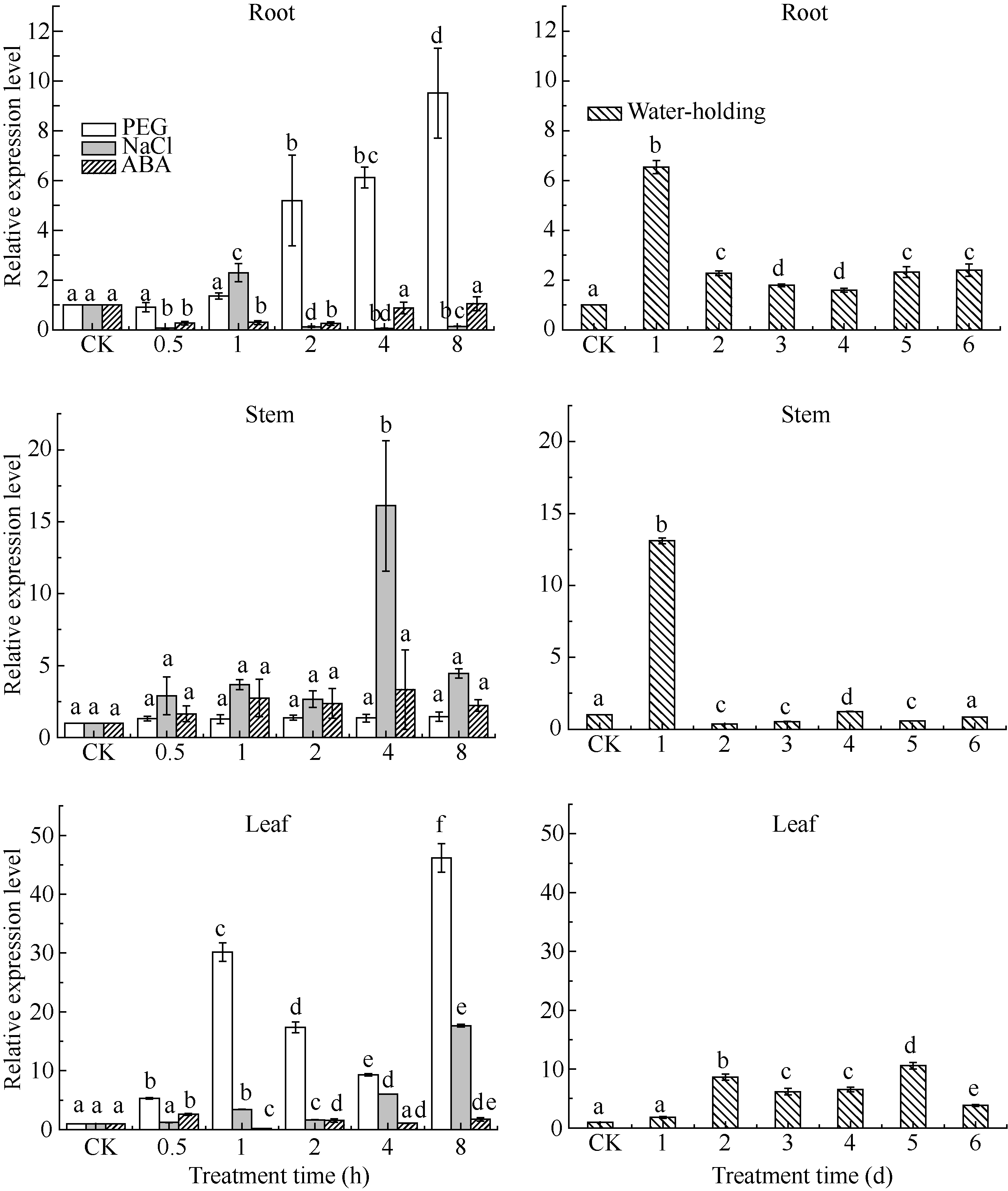
图6 SiCDPK1基因表达模式分析柱上小写字母不同表示在0.05的水平上差异显著。Fig. 6 Expression pattern analysis of SiCDPK1 geneBars superscripted by different letters are significantly different at 0.05 probability level.
BoudsocqM, SheenJ. CDPKs in immune and stress signaling. Trends Plant Sci, 2013, 18: 30-40[本文引用:4][JCR: 11.808]
[2]
RutschmannF, StalderU, PiotrowskiM, OeckinqC, SchallerA. LeCPK1, a calcium-dependent protein kinase from tomato. Plasma membrane targeting and biochemical characterization. Plant Physiol, 2002, 129: 156-168[本文引用:3][JCR: 6.555]
[3]
LuS X, HrabakE M. An Arabidopsis calcium-dependent protein kinase is associated with the endoplasmic reticulum. Plant Physiol, 2002, 128: 1008-1021[本文引用:2][JCR: 6.555]
[4]
MartinM L, BusconiL. Membrane localization of a rice calcium-dependent protein kinase (CDPK) is mediated by myristoylation and palmitoylation. Plant J, 2000, 24: 429-435[本文引用:2][JCR: 6.582]
[5]
BenetkaW, MehlmerN, Maurer-StrohS, SammerM, Korand aM, NeumüllerR, BetschinqerJ, KnoblichJ A, TeiqeM, EisenhaberF. Experimental testing of predicated myristoylation targets involved in asymmetric cell division and calcium-dependent signaling. Cell Cycle, 2008, 7: 3709-3719[本文引用:2][JCR: 5.243]
[6]
ChehabE W, PatharkarO R, HeqemanA D, TaybiT, CushmanJ C. Autophosphorylation and subcellular localization dynamics of a salt- and water defict-induced calcium-dependent protein kinase from ice plant. Plant Physiol, 2004, 135: 1430-1446[本文引用:1][JCR: 6.555]
[7]
MehlmerN, WurzinqerB, StaelS, Hofmann-RodriquesD, CsaszarE, PfisterB, BayerR, TeiqeM. The Ca2+-dependent protein kinase CPK3 is required for MAPK-independent salt-stress acclimation in Arabidopsis. Plant J, 2010, 63: 484-498[本文引用:2][JCR: 6.582]
[8]
StaelS, BayerR G, MehlmerN, TeiqeM. Protein N-acylation overrides differing targeting signals. FEBS Lett, 2011, 585: 517-522[本文引用:1][JCR: 3.582]
[9]
WitteC P, KeinathN, DubiellaU, DemoulièreR, SealA, RomeisT. Tobacco calcium-dependent protein kinases are differentially phosphorylated in vivo as part of a kinase cascade that regulates stress response. J Biol Chem, 2010, 285: 9740-9748[本文引用:1][JCR: 4.651]
[10]
ChristodoulouJ, MalmendalA, HarperJ F, ChazinW J. Evidence for differing roles for each lobe of the calmodulin-like domain in a calcium-dependent protein kinase. J Biol Chem, 2004, 279: 29092-29100[本文引用:1][JCR: 4.651]
[11]
HarperJ F, BretonG, HarmonA. Decoding Ca2+ signals through plant protein kinases. Annu Rev Plant Biol, 2004, 55: 263-288[本文引用:1][JCR: 25.962]
[12]
HarperJ F, HarmonA. Plants, symbiosis and parasites: a calcium signaling connection. Nat Rev Mol Cell Biol, 2005, 6: 555-566[本文引用:1]
[13]
ChengS H, WillmannM R, ChenH C, SheenJ. Calcium signaling through protein kinases. The Arabidopsis calcium-dependent protein kinase gene family. Plant Physiol, 2002, 129: 469-485[本文引用:4][JCR: 6.555]
[14]
AsanoT, TanakaN, YangG, HayashiN, KomatsuS. Genome-wide identification of the rice calcium-dependent protein kinase and its closely related kinase gene families: comprehensive analysis of the CDPKs gene family in rice. Plant Cell Physiol, 2005, 46: 356-366[本文引用:1][JCR: 4.134]
[15]
RayS, AqarwalP, AroraR, KapoorS, TyaqiA K. Expression analysis of calcium-dependent protein kinase gene family during reproductive development and abiotic stress conditions in rice (Oryza sativa L. ssp. indica). Mol Genet Genomics, 2007, 278: 493-505[本文引用:2][JCR: 2.881]
[16]
LiA L, ZhuY F, TanX M, WangX, WeiB, GuoH Z, ZhangZ L, ChenX B, ZhaoG Y, KongX Y, JiaJ Z, MaoL. Evolutionary and functional study of the CDPK gene family in wheat (Triticum aestivum L. ). Plant Mol Biol, 2008, 66: 429-443[本文引用:3][JCR: 3.518]
[17]
MaP, LiuJ, YangX, MaR. Genome-wide identification of the maize calcium-dependent protein kinase gene family. Appl Biochem Biotechnol, 2013, 169: 2111-2125[本文引用:1][JCR: 1.348]
[18]
MyersC, RomanowskyS M, BarronY D, GarqS, AzuseC L, CurranA, DavisR M, HattonJ, HarmonA C, HarperJ F. Calcium-dependent protein kinase regulate polarized tip growth in pollen tubes. Plant J, 2009, 59: 528-539[本文引用:2][JCR: 6.582]
ZouJ J, WeiF J, WangC, WuJ J, RatnasekeraD, LiuW X, WuW H. Arabidopsis calcium-dependent protein kinase CPK10 functions in abscisic acid- and Ca2+-mediated stomatal regulation in response to drought stress. Plant Physiol, 2010, 154: 1232-1243[本文引用:1][JCR: 6.555]
[21]
YuX C, LiM J, GaoG F, FengH Z, GengX Q, PengC C, ZhuS Y, WangX J, ShenY Y, ZhangD P. Abscisic acid stimulates a calcium-dependent protein kinase in grape berry. Plant Physiol, 2006, 140: 558-579[本文引用:1][JCR: 6.555]
[22]
ZhuS Y, YuX C, WangX J, ZhaoR, LiY, FanR C, ShangY, DuS Y, WangX F, WuF Q, XuY H, ZhangX Y, ZhangD P. Two calcium-dependent protein kinases, CPK4 and CPK11, regulate abscisic acid signal transduction in Arabidopsis. Plant Cell, 2007, 19: 3019-3036[本文引用:1][JCR: 9.251]
[23]
DammannC, IchidaA, HonqB, RomanowskyS M, HrabakE M, HarmonA C, PickardB G, HarperJ F. Subcellular targeting of nine calcium-dependent protein kinase isoforms from Arabidopsis. Plant Physiol, 2003, 132: 1840-1848[本文引用:1][JCR: 6.555]
[24]
ChoiH I, ParkH J, ParkJ H, KimS, ImM Y, SeoH H, KimY W, HwangI, KimS Y. Arabidopsis calcium-dependent protein kinase AtCPK32 interacts with ABF4, a transcriptional regulator of abscisic acid-responsive gene expression, and modulates its activity. Plant Physiol, 2005, 139: 1750-1761[本文引用:2][JCR: 6.555]
[25]
CocaM, SanSegundo B. AtCPK1 calcium-dependent protein kinase mediates pathogen resistance in Arabidopsis. Plant J, 2010, 63: 526-540[本文引用:1][JCR: 6.582]
[26]
HarperJ F, SussmanM R, SchallerG E, Putnam-EvansC, CharbonneauH, HarmonA C. A calcium-dependent protein kinase with a regulatory domain similar to calmodulin. Science, 1991, 252: 951-954[本文引用:1]
[27]
RomeisT, LudwigA A, MartinR, Jones J D G. Calcium- dependent protein kinases play an essential role in a plant defence response. EMBO J, 2001, 20: 5556-5567[本文引用:1][JCR: 9.822]
[28]
WanB, LinY, MouT. Expression of rice Ca2+-dependent protein kinases (CDPKs) genes under different environmental stresses. FEBS Lett, 2007, 581: 1179-1189[本文引用:2][JCR: 3.582]
[29]
KanchiswamyC N, TakahashiH, QuadroS, MaffeiM E, BossiS, BerteaC, ZebeloA S, MuroiA, IshihamaN, YoshiokaH, Boland W, TakabayashiJ, EndoY, SawasakiT, ArimuraG. Regulation of Arabidopsis defense responses against Spodoptera littoralis by CPK-mediated calcium signaling. BMC Plant Biol, 2010, 10-97[本文引用:1][JCR: 4.354]
[30]
IshidaS, YuasaT, NakataM, TakahashiY. A tobacco calcium- dependent protein kinase, CDPK1, regulates the transcription factor REPRESSION OF SHOOT GROWTH in response to gibberellins. Plant Cell, 2008, 20: 3273-3288[本文引用:1][JCR: 9.251]
[31]
ItoT, NakataM, FukazawaJ, IshidaS, TakahashiY. Alteration of substrate specificity: the variable N-terminal domain of tobacco Ca2+-dependent protein kinase is important for substrate recognition. Plant Cell, 2010, 22: 1592-1604[本文引用:1][JCR: 9.251]
[32]
SchulzP, HerdeM, RomeisT. Calcium-dependent protein kinases: hubs in plant stress signaling and development. Plant Physiol, 2013, 163: 523-530[本文引用:2][JCR: 6.555]
[33]
HubbardK E, SieqelR S, ValerioG, Brand tB, SchroederJ I. Absicsic acid and CO2 signalling via calcium sensitivity priming in guard cells, new CDPK mutant phenotypes and a method for improved resolution of stomatal stimulus-response analysis. Ann Bot, 2012, 109: 5-17[本文引用:2][JCR: 0.657]
[34]
HarmonA C, GribskovM, HarperJ F. CDPKs-a kinase for every Ca2+ signal?Trends Plant Sci, 2000, 5: 154-159[本文引用:2][JCR: 11.808]
[35]
LieseA, RomeisT. Biochemical regulation of in vivo function of plant calcium-dependent protein kinase (CDPK). Biochim Biophy Acta, 2013, 1833: 1582-1589[本文引用:3]
[36]
HrabakE M, ChenC W, GribskovM, HarperJ F, ChoiJ H, HalfordN, KudlaJ, LuanS, NimmoH G, SussmanM R, ThomasM, Walker-SimmonsK, ZhuJ K, HarmonA C. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol, 2003, 132: 666-680[本文引用:2][JCR: 6.555]
[37]
LudwiqA A, RomeisT, JonesJ D. CDPK-mediated signaling pathways: specificity and cross-talk. J Exp Bot, 2004, 55: 181-188[本文引用:2][JCR: 5.242]
[38]
LataC, GuptaS, PrasadM. Foxtail millet: a model crop for genetic and genomic studies in bioenergy grasses. Crit Rev Biotechnol, 2013, 33: 328-343[本文引用:2][JCR: 5.095]
[39]
李志江. 谷子抗除草剂基因的发现及其应用. 基因组学与应用生物学, 2010, 29: 768-774LiZ J. Discovery and application of herbicide resisitant gene in foxtail millet. Genom Appl Biol, 2010, 29: 768-774 (in Chinese with English abstract)[本文引用:1][CJCR: 0.7323]
[40]
瓮巧云, 宋晋辉, 张爱香. 谷子丝/苏氨酸蛋白激酶类抗病基因同源序列的克隆与分析. 河南农业科学, 2012, 41: 106-108WengQ Y, SongJ H, ZhangA X. Cloning and analysis of STK disease resistant gene analogs in millet. J Henan Agric Sci, 2012, 41: 106-108 (in Chinese with English abstract)[本文引用:1]
[41]
崔润丽, 智慧, 王永芳, 李伟, 李海权, 黄占景, 刁现民. 谷子DnaJ蛋白基因的克隆. 华北农学报, 2007, 22(4): 9-13CuiR L, ZhiH, WangY F, LiW, LiH Q, HuangZ J, DiaoX M. Cloning of DnaJ-like protein gene from foxtail millet. Acta Agric Boreali-Sin, 2007, 22(4): 9-13 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
[42]
杨希文, 胡银岗. 谷子DREB转录因子基因的克隆及其在干旱胁迫下的表达模式分析. 干旱地区农业研究. 2011, 29(5): 69-74YangX W, HuY G. Cloning of a DREB gene from foxtail millet (Setaria italica L) and its expression during drought stress. Agric Res Arid Areas, 2011, 29(5): 69-74 (in Chinese with English abstract)[本文引用:2]
[43]
ZhangJ P, ZhengJ, ZhuY, GuoJ F, WangG Y. Cloning and characterization of a putative 12-oxophytodienoic acid reductase cDNA induced by osmotic stress in roots of foxtail millet. DNA Seq, 2007, 18: 138-144[本文引用:1]
[44]
崔润丽, 智慧, 王永芳, 李伟, 李海权, 黄占景, 刁现民. 谷子3-磷酸甘油醛脱氢酶基因的克隆与结构分析. 华北农学报, 2009, 24(3): 10-14CuiR L, ZhiH, WangY F, LiW, LiH Q, HuangZ J, DiaoX M. Cloning and structure analysis of Foxtail Millet APDH gene. Acta Agric Boreali-Sin, 2009, 24(3): 10-14 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
[45]
PengY L, ZhangJ P, CaoG Y. Overexpression of a PLDα1 gene from Setaria italica enhances the sensitivity of Arabidopsis to abscisic acid and improves its drought tolerance. Plant Cell Rep, 2010, 29: 793-802[本文引用:1][JCR: 2.509]
[46]
赵晋锋, 余爱丽, 田岗, 杜艳伟, 郭二虎, 刁现民. 谷子CBL基因鉴定及其在干旱、高盐胁迫下的表达分析. 作物学报, 2013, 39: 360-367ZhaoJ F, YuA L, TianG, DuY W, GuoE H, DiaoX M. Identification of CBL genes from foxtail millet (Setaria italica [L. ] Beauv. ) and its expression under drought and salt stresses. Acta Agron Sin, 2013, 39: 360-367 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[47]
张雁明, 王莉, 张彬, 王海岗, 彭锁堂, 李萍, 韩渊怀. 谷子ABF3基因对PEG胁迫的响应. 山西农业大学学报(自然科学版), 2013, 33: 191-196ZhangY M, WangL, ZhangB, WangH G, PengS T, LIP, HanY H. The response of ABF3 Gene to PEG stress in foxtail millet (Setaria italica L. ). J Shanxi Agric Univ (Nat Sci Edn), 2013, 33: 191-196[本文引用:1]
[48]
李志江, 刁现民. 谷子分子标记与功能基因组研究进展. 中国农业科技导报, 2009, 11(4): 16-22LiZ J, DiaoX M. Research progress on molecular marker and functional genomic of foxtail millet (Setaria italic Beauv. ). J Agric Sci Technol, 2009, 11(4): 16-22 (in Chinese with English abstract)[本文引用:1]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}