 , 王新超
, 王新超*通讯作者(Corresponding authors): 王新超, E-mail:xcw75@tricaas.com, Tel: 0571-86653162; 杨亚军, E-mail:yjyang@tricaas.com, Tel: 0571-86650226 收稿日期:2014-04-16 基金:
摘要
关键词:茶树; bZIP转录因子;
Molecular Cloning and Expression of a bZIP Transcription Factor Gene
CAO Hong-Li, YUE Chuan, ZHOU Yan-Hua, WANG Lu, HAO Xin-Yuan, YANG Ya-Jun
, WANG Xin-ChaoFund:
Abstract
Keyword:Tea plant (
Show Figures
Show Figures





茶树( Camellia sinensis)原产于热带及亚热带, 是一种喜温畏寒的经济作物。茶树生长和茶叶生产受低温、干旱等环境条件的制约, 发掘茶树抗性基因, 培育抗性较强的茶树品种有重大意义。转录因子(transcription factors, TFs)在植物响应逆境胁迫中具有重要的作用。它能够激活或者抑制下游基因的转录, 使植物适应或抵御外界环境的变化。近年来, AP2/EREBP、MYB、NAC、WRKY和bZIP等转录因子在植物逆境响应中被广泛研究[ 1]。其中, bZIP是真核生物中分布最广、最保守的一类转录因子[ 2], 与植物抵御多种逆境胁迫密切相关, 但在茶树上还没有相关研究报道。因此, 克隆茶树bZIP转录因子基因, 分析其不同逆境胁迫下的表达特性, 为茶树抗性机制研究与抗性育种提供理论基础有重要意义。
bZIP转录因子(basic region/leucine zipper proetin), 由一个与DNA结合的碱性结构域和一个亮氨酸拉链二聚体组成, 是转录因子中最大家族之一[ 2]。植物bZIP蛋白主要专一结合核心序列为ACGT回文结构的顺式作用元件, 如A-box (TACGTA)、C-box (GACGTC)和G-box (CACGTG)等元件[ 3]。目前已经在拟南芥[ 4]、水稻[ 5]和豆科植物[ 6]等基因组中发现大量的bZIP家族基因。它们参与多种生物学过程, 包括光信号传导、花的发育、种子成熟、病菌防御以及各种非生物胁迫的响应等[ 4, 7, 8, 9, 10]。近年来, 已从玉米[ 11]、番茄[ 12]、高粱[ 13]、胡椒[ 14]、苎麻[ 15]等植物中克隆到逆境相关的bZIP转录因子, 并证实它们与植物抗性密切相关。Wang等[ 16]在烟草中过表达刚毛柽柳( Tamarix hispida) ThbZIP1, 能显著提高转基因烟草的耐盐性。Chen等[ 17]在水稻中克隆到一个 OsbZIP16, 进一步研究显示 OsbZIP16能正调控水稻抗旱性, 并且转基因水稻的抗寒性显著增强。Liao等[ 8]在拟南芥中过表达水稻bZIP转录因子基因 GmbZIP44、 GmbZIP46、 GmbZIP62和 GmbZIP78, 结果显示转基因植株的耐盐性和抗寒性都得到提高。
前期, 本课题组进行了茶树冷驯化转录组测序, 从中获得了一批受低温诱导差异表达的bZIP转录因子的EST片段[ 18]。本研究通过RACE技术, 克隆茶树 bZIP基因的cDNA全长, 并对其进行生物信息学分析和亚细胞定位, 以实时荧光定量PCR分析其在低温、ABA和高盐胁迫处理下的表达模式, 探究bZIP转录因子与茶树抗逆性的关系, 为茶树抗逆基因工程提供候选基因资源和理论依据。
1 材料与方法1.1 材料和试剂所用的茶树品种为特早生国家级茶树品种龙井43, 种植于中国农业科学院茶叶研究所国家种质杭州茶树圃。参照已报道的茶树胁迫处理[ 19, 20, 21]及本实验室前期试验结果[ 22], 从日光温室中选取生长正常且长势相近的3年生盆栽茶树若干盆, 分别在低温(4℃)、ABA (100 μmol L-1)和NaCl (250 mmol L-1)胁迫下试验。
SMART RACE cDNA Amplication Kit和Advantage Polymerase Mix购于Clontech公司(Mountain View, CA, USA); SYBR Premix Ex Taq II (Perfect Real Time)、DNA Marker、TA Cloning Kit PCR2.1 Vector、大肠杆菌DH5α感受态细胞和 Taq聚合酶为TaKaRa公司产品(大连); DNase I Digestion of RNA Preparation、First-Strand cDNA Synthesis购自Invitrogen公司(Carlsbad, USA); 琼脂糖凝胶回收试剂盒购自Axygen公司(California, USA); RNA提取试剂盒购自天根公司(Tiangen, 北京)。脱落酸(ABA)购于SIGMA公司, NaCl、无水乙醇等为国产分析纯试剂。由上海华津生物技术公司合成引物并测序。
1.2 总RNA提取和cDNA合成参照离心柱型植物总RNA快速提取试剂盒说明书提取茶树叶片总RNA。检测RNA浓度和完整性后于-80℃保存备用。用1 μg总RNA为模板, 参照SMART RACE cDNA Synthesis Kit的操作步骤合成5′和3′的cDNA, 用于RACE扩增, 以及反转录PCR (RT-PCR)扩增基因的全长开放阅读框(ORF)和亚细胞定位载体构建的PCR扩增; 以5 μg总RNA为模板, 按照DNase I Digestion of RNA Preparation和First-Strand cDNA Synthesis试剂盒说明书合成cDNA, 稀释40倍后作实时荧光定量PCR (qRT-PCR)模板, 进行qRT-PCR检测。
1.3 CsbZIP1全长cDNA克隆与生物信息学分析 1.3.1 茶树 CsbZIP1基因的克隆 根据本实验室茶树冷驯化转录组测序结果(NCBI登录号为SRA061043), 经Blast比对筛选出与其他植物中报道的 bZIP基因高度同源的EST序列, 设计5′和3′-RACE特异引物5′GSP/3′GSP (表1)。参照SMART RACE cDNA Synthesis Kit的操作步骤进行茶树 bZIP基因的PCR反应, 反应条件为94℃预变性4 min; 94℃变性30 s, 68℃退火30 s, 72℃延伸2 min, 30个循环; 72℃延伸10 min。产物经1%琼脂糖凝胶电泳检测, 将目的条带回收纯化, 连接到PCR2.1载体, 转化大肠杆菌DH5α, 蓝白斑筛选阳性菌落, 挑取10个白色单菌落进行菌落PCR检测, 挑选3~4个阳性菌落送上海华津生物公司测序。经去载体、拼接, 得到茶树 bZIP基因的5′和3′端序列, 与中间序列拼接, 得到 bZIP基因的全长cDNA序列。在ORF上、下游区域设计RT-PCR特异引物RT-F/RT-R (表1), 以PCR扩增验证ORF序列。反应条件为94℃ 4 min; 94℃ 30 s, 58℃ 30 s, 72℃ 1 min, 35个循环。PCR产物经回收、连接、转化、测序和拼接后, 获得完整的茶树 bZIP基因全长cDNA序列。
表1
Table 1
表1(Table 1)
| 表1 试验引物及其序列 Table 1 Sequence of primers in this study |
1.3.2 CsbZIP1生物信息学分析 利用NCBI的BlastX和BlastP进行核苷酸和氨基酸序列比对分析; 用DNAStar 软件查询开放阅读框; 用ProtParam预测蛋白质分子量和理论等电点; 用SMART (http://smart.embl-heidelberg.de/)进行保守域分析; 用DNAMAN进行多序列氨基酸比对; 用MEGA5.0软件中的邻近相连法构建系统进化树; 用NetPhos 2.0 Serve (http://www.cbs.dtu.dk/services/NetPhos/)预测蛋白质磷酸化位点; 用SWISS-MODEL (http://swissmodel.expasy.org/workspace/)和PyMol软件预测蛋白质三级结构; 用WoLF PSORT (http://wolfpsort.org/)进行亚细胞定位预测。
1.4 CsbZIP1基因编码蛋白的亚细胞定位 1.4.1 载体的构建 以CsbZIP1-GFP-F/CsbZIP1-GFP- R为引物, 扩增 CsbZIP1基因, 纯化扩增产物, 用 Kpn I和 Xho I酶切, 回收酶切产物; 再用 Kpn I和 Xho I对P2GWF7质粒进行酶切反应; 最后, 把酶切回收的PCR产物和质粒酶切片段连接, 构建载体p2GWF7-CsbZIP1, 转入DH5α, 菌液PCR、酶切筛选阳性克隆, 并对阳性克隆进行测序验证。
1.4.2 P2GWF7-CsbZIP1转化洋葱表皮细胞 取 50 mg mL-1金粉悬液50 μL于离心管, 依次加入5 μL质粒DNA (1 μg μL-1), 50 μL CaCl2(2.5 mol L-1)和20 μL亚精胺(0.1 mol L-1)(每加完一样可振荡3 s), 静置10 min; 然后振荡1 min, 冰上10 min, 9400× g离心10 s, 弃上清; 再用80 μL无水乙醇, 洗涤沉淀一次; 重新悬浮沉淀于10 μL无水乙醇中; 每次用10 μL滴于微粒载膜中央, 平铺, 晾至完全干燥, 待用。取新鲜洋葱, 用无菌刀片切取肉质较厚的3~4层鳞茎, 用镊子撕出表皮平铺于MS培养基上, 25℃预培养4 h。选用1100 psi的压力膜, 将10 μL金粉DNA混合物滴在压力膜中央, 将洋葱表皮放在琼脂糖培养基上, 采用PDS1000/He型基因枪(Bio-Rad)轰击。将轰击后的洋葱表皮细胞25℃暗培养16 h后制片, 于激光共聚焦显微镜下观察并照相。
1.5 CsbZIP1实时荧光定量PCR表达分析 以茶树 18S基因作为内参基因, 参照SYBR Premix Ex Taq II (Perfect real-time)的操作说明书对 CsbZIP1基因的表达进行荧光定量PCR分析, PCR体系为SYBR Premix Ex Taq25 μL、10 μmol L-1上下游引物各1 μL、ROX Dye II 1 μL、cDNA 2 μL, 加水至终体积50 μL。反应在ABI PRISM 7500实时定量PCR仪上进行, 程序为95℃ 15 s, 94℃ 5 s, 60℃ 34 s, 40个循环。反应结束后分析荧光值变化曲线和熔解曲线, 采用2-ΔΔCT法分析结果[ 23], 每个样品3次重复。
2 结果与分析2.1 CsbZIP1基因的克隆及生物信息学分析 2.1.1 CsbZIP1全长cDNA克隆与序列分析 根据中间EST片段序列, 设计5′和3′-RACE特异引物, 分别扩增出928 bp和410 bp的序列(图1), 用SeqMan软件把3段序列拼接初步得到cDNA全长, 再查找其开放阅读框(ORF), BlastX比对其ORF, 在ORF上下游设计引物进行RT-PCR扩增验证, 扩增得到813 bp的序列(图1)。最终获得该基因cDNA全长序列1515 bp, 包含813 bp的完整ORF, 编码270个氨基酸, 理论等电点为7.175, 推测分子量29.484 kD。NCBI比对分析结果表明, 它含有bZIP家族典型的BRLZ结构域, 属于B-zip1家族; 与可可( Theobroma cacao) EOX97663.1、大豆( Glycine max) NP-001235033.1的氨基酸相似性分别为68%和66%。CsbZIP1蛋白与其他植物bZIP氨基酸序列进行多序列比对表明, 在N端具有保守的bZIP碱性结构域和亮氨酸拉链(图2)。对CsbZIP1氨基酸序列分析, 显示氨基酸序列
图1
Fig. 1
| Figure OptionViewDownloadNew Window | |
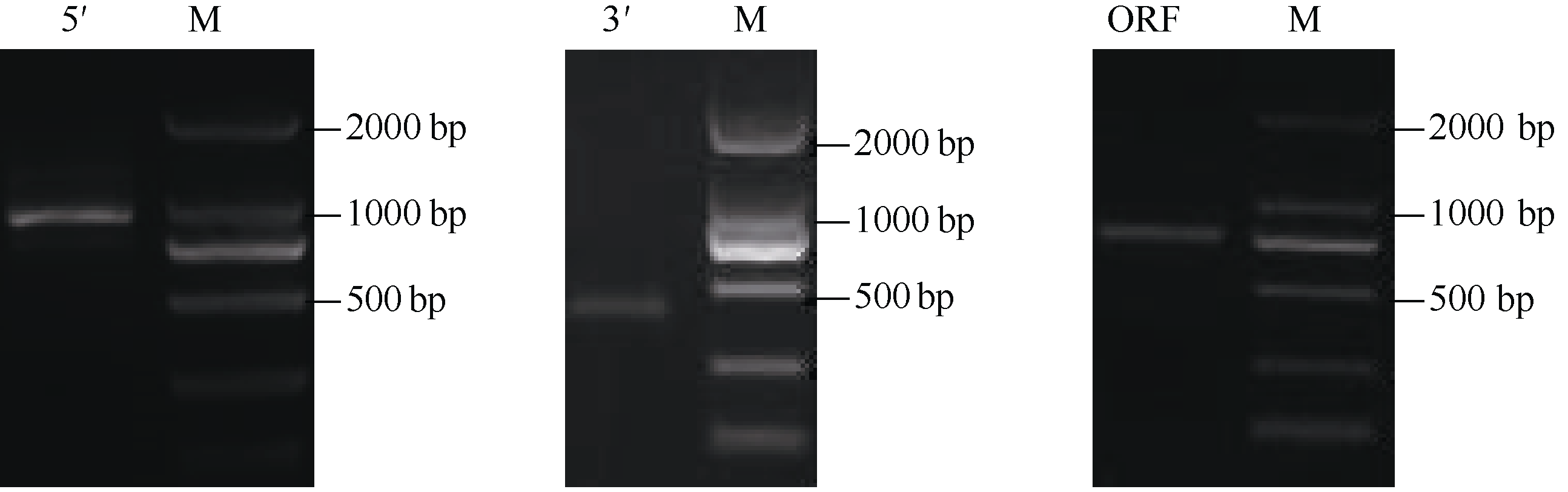 | 图1 CsbZIP1的RACE及RT-PCR电泳结果Fig. 1 Electrophoresis results of RACE fragments and RT-PCR fragments of CsbZIP1 |
图2
Fig. 2
| Figure OptionViewDownloadNew Window | |
 | 图2 CsbZIP1与其他植物同源bZIP的氨基酸多序列比对黑色画线标记为bZIP家族保守的N-x7-R/K-x9-L-x6-L-x6-L序列。Fig. 2 Multiple amino acid alignment of CsbZIP1 with other species homologous bZIPConserved sequence N-x7-R/K-x9-L-x6-L-x6-L of bZIP family is underlined. |
第95~127位为典型的bZIP碱性结构域和亮氨酸拉链结构域(图3)。因此, 该基因属于bZIP转录因子家族, 并命名为 CsbZIP1 (GenBank登录号为JX050148.1)。
图3
Fig. 3
| Figure OptionViewDownloadNew Window | |
 | 图3 CsbZIP1的cDNA核苷酸序列及预测氨基酸序列图中阴影部分为碱性结构域(N-x7-R/K-x9); 下画线部分为亮氨酸拉链结构域(L-x6-L-x6-L), 其中黑色反白标识为亮氨酸(Leucine)。Fig. 3 Nucleotide and putative amino acid sequence of CsbZIP1The shaded parts are basic region (N-x7-R/K-x9); underlined parts are leucine zipper motif; and black parts are leucine. |
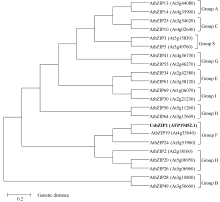
2.1.2 CsbZIP1蛋白与拟南芥bZIP蛋白的系统进化树分析 根据Jakoby等[ 4]对拟南芥bZIP亚家族的分类, 从每个亚家族挑选取2~3个拟南芥 bZIP基因, 采用MEGA 5.0邻近相连法对CsbZIP1与21个拟南芥bZIP氨基酸序列比对, 并构建系统进化树(图4), 结果显示CsbZIP1与拟南芥Group F聚为一类, 从理论上推测CsbZIP1属于bZIP转录因子的F亚家族。
图4
Fig. 4
| Figure OptionViewDownloadNew Window | |
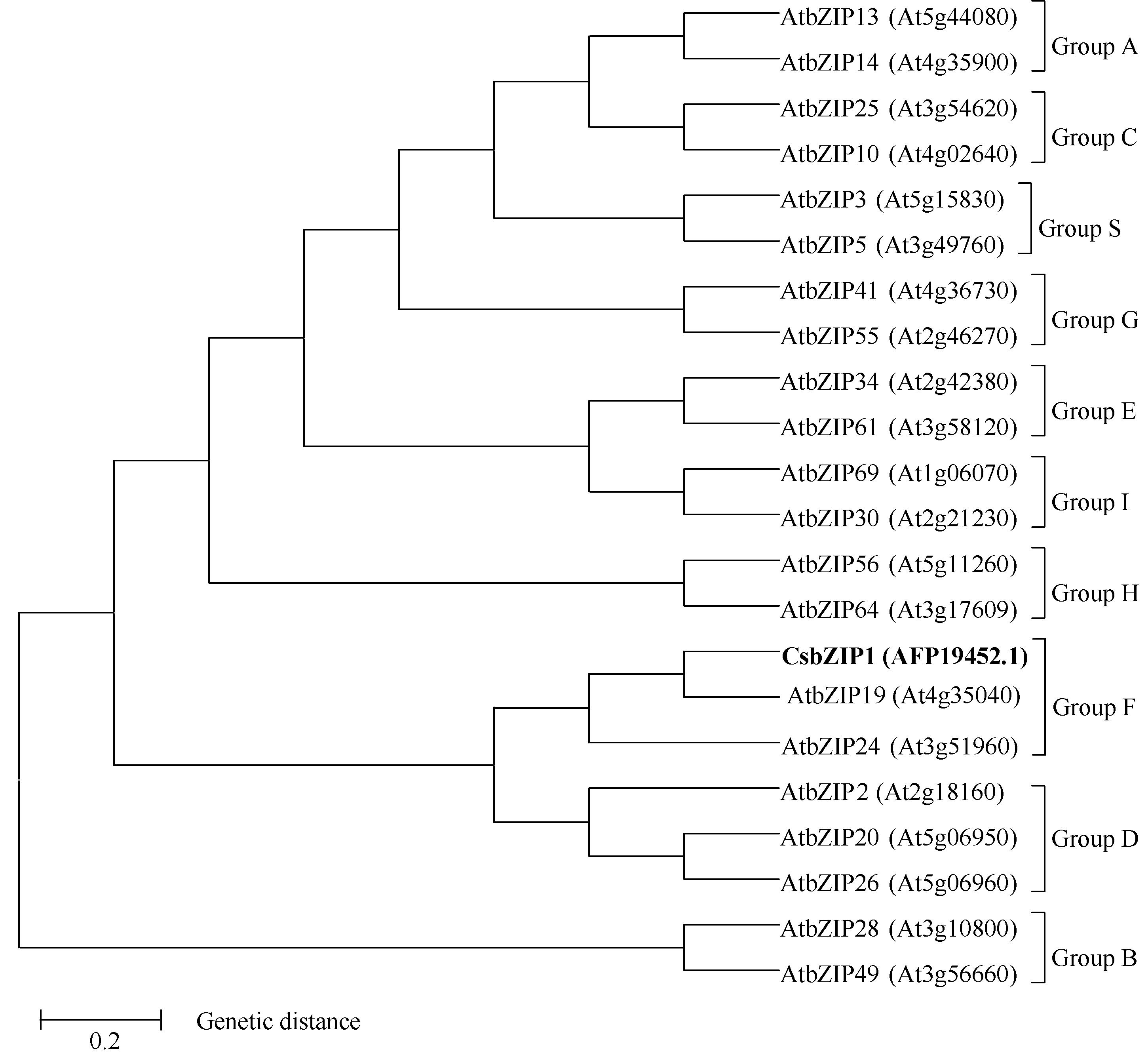 | 图4 CsbZIP1蛋白与21个拟南芥bZIP转录因子蛋白的进化树分析Fig. 4 Phylogenetic tree analysis of CsbZIP1 protein with 21 Arabidopsis bZIP proteins |
2.1.3 CsbZIP1编码蛋白的磷酸化位点预测 用NetPhos 2.0 Serve预测CsbZIP1蛋白磷酸化位点(图5)表明, 含有丝氨酸(Serine)、苏氨酸(Threonine)和酪氨酸(Tyrosine)的多个磷酸化位点中以丝氨酸位点最多, 有13个, 苏氨酸和酪氨酸位点各1个。由此推测磷酸化作用可能与CsbZIP1蛋白的活性调控有关。
图5
Fig. 5
| Figure OptionViewDownloadNew Window | |
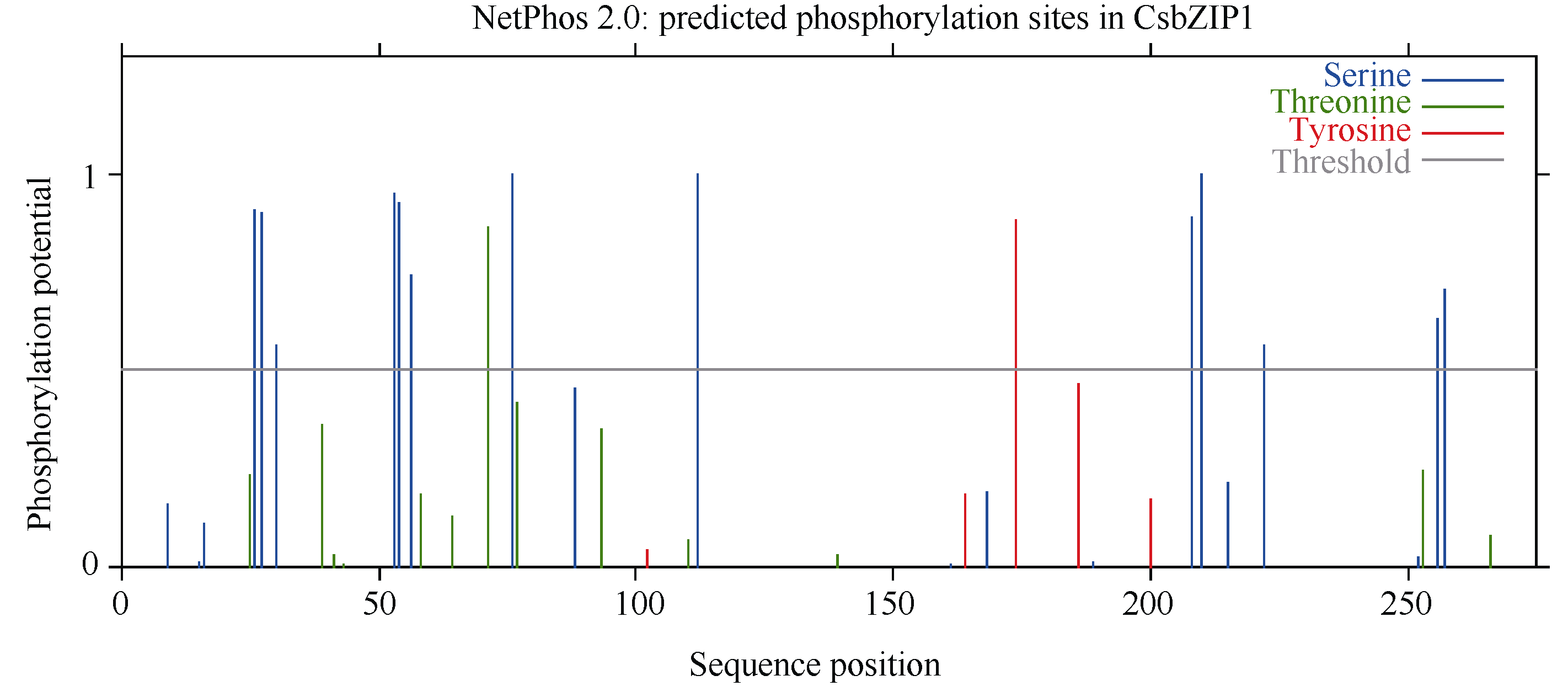 | 图5 CsbZIP1蛋白磷酸化位点预测Fig. 5 Phosphorylation site prediction of CsbZIP1 protein |
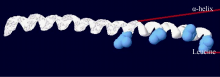
2.1.4 CsbZIP1编码蛋白的三级结构预测 用SWISS- MODEL预测CsbZIP1蛋白的三级结构, 经过PyMol软件模拟分析, 得到CsbZIP1蛋白的三维结构图(图6)。结果显示与Jakoby等[ 4]获得的拟南芥bZIP蛋白的三级结构模型图相似, 均含有bZIP转录因子典型的α-螺旋和亮氨酸拉链结构, 进一步证实获得的 CsbZIP1属于bZIP转录因子家族。
图6
Fig. 6
| Figure OptionViewDownloadNew Window | |
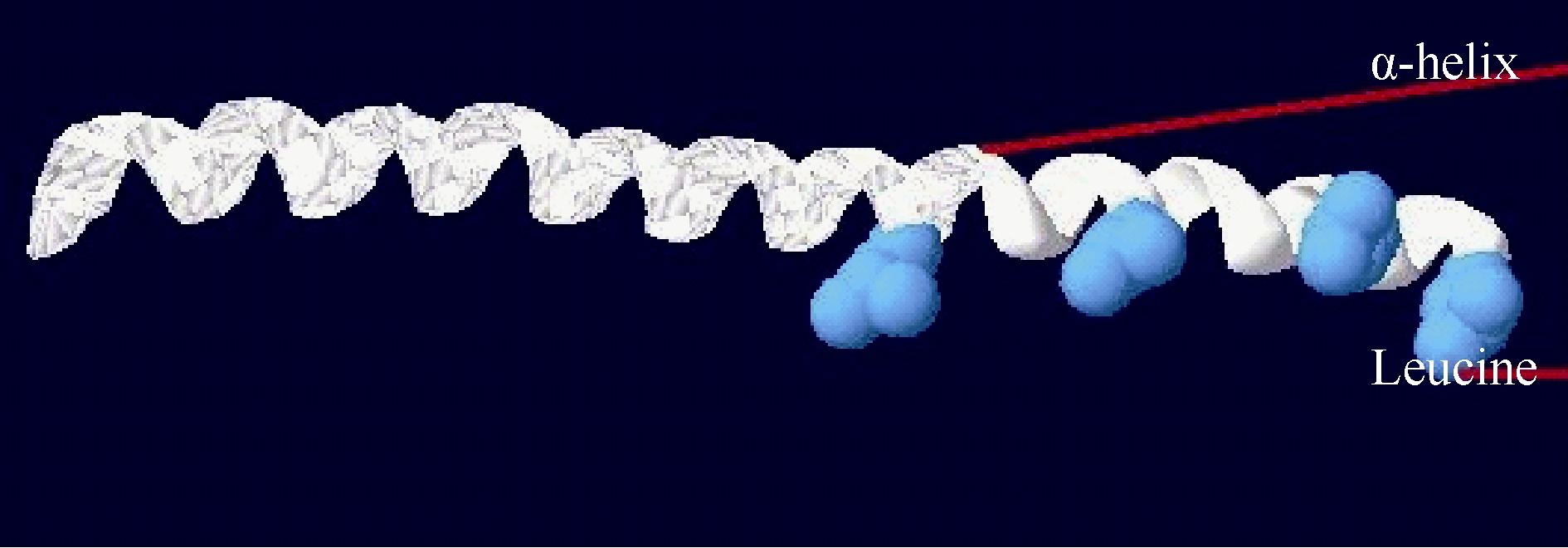 | 图6 CsbZIP1蛋白三维结构图Fig. 6 3-D structure of CsbZIP1 protein |
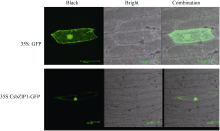
2.2 CsbZIP1编码蛋白的亚细胞定位用WoLF PSORT软件进行亚细胞定位预测显示, CsbZIP1主要定位在细胞核内。洋葱表皮亚细胞定位试验的结果也显示CsbZIP1融合蛋白位主要定位于细胞核内(图7), 但是细胞内其他部位也检测到微弱信号。bZIP定位在细胞核内与它的功能密切相关。
图7
Fig. 7
| Figure OptionViewDownloadNew Window | |
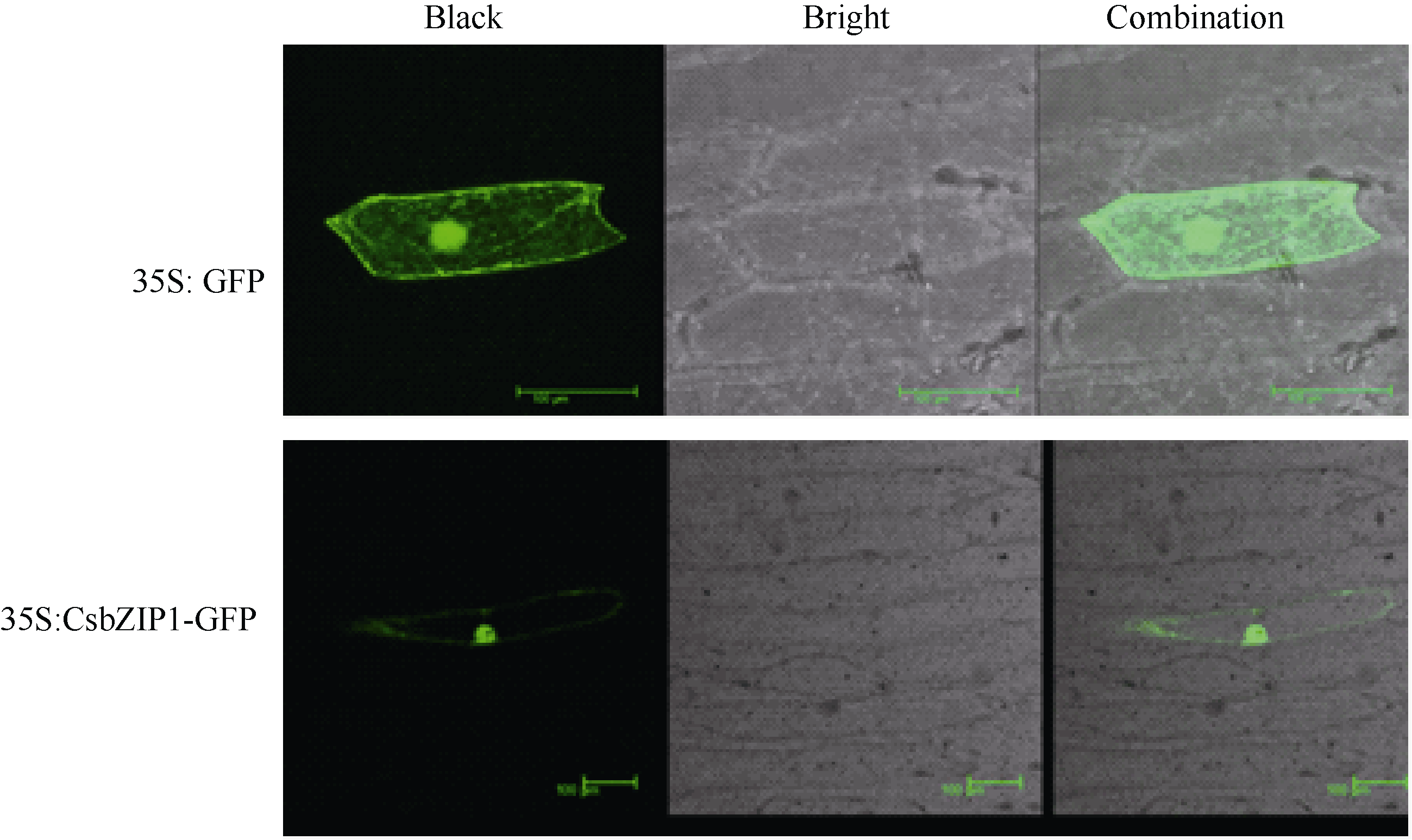 | 图7 CsbZIP1亚细胞定位Fig. 7 Subcellular localization of CsbZIP1 |
2.3 CsbZIP1在低温、ABA和NaCl胁迫处理下的实时荧光定量分析 荧光定量PCR结果表明(图8), 4℃低温处理能够诱导 CsbZIP1的表达, 且随着处理时间的延长, 表达量在处理 8 h最大, 达到11.59, 24 h降低为6.72。100 μmol L-1ABA 处理后 CsbZIP1的表达被抑制, 2 h的相对表达量最低只有0.2, 但随着胁迫的继续, 表达量逐渐上升, 在24 h时表达量升高到0.90, 但没有超过0 h的表达量。250 mmol L-1NaCl处理后, CsbZIP1的表达量呈先升高后降低的趋势, 处理4 h表达被诱导, 表达量达3.86, 随着处理时间延长至24 h, 表达量降低为1.21。 CsbZIP1在低温、高盐和ABA胁迫下均响应, 低温能够显著诱导其表达, NaCl短时间内也能上调其表达, ABA则能迅速抑制其表达。
图8
Fig. 8
| Figure OptionViewDownloadNew Window | |
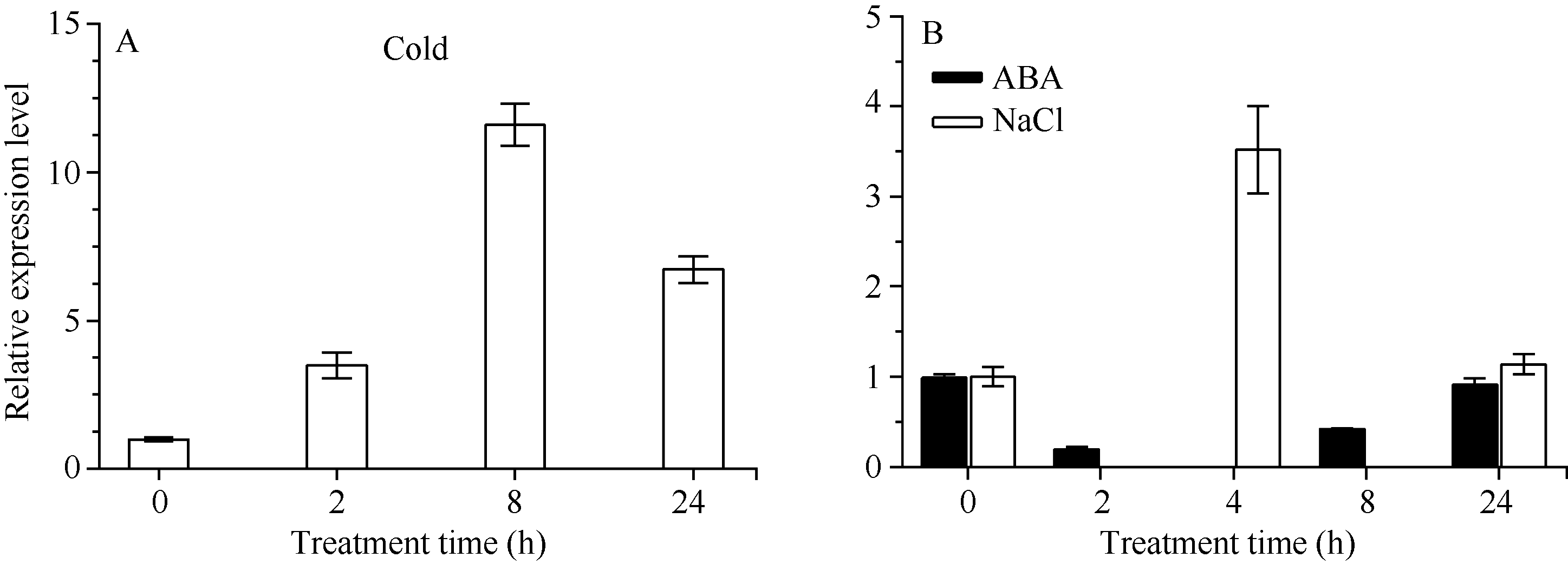 | 图8 逆境胁迫处理下 CsbZIP1的相对表达量Fig. 8 Relative expression of CsbZIP1under stress treatments |
3 讨论bZIP作为一类重要的转录因子, 与植物逆境胁迫响应密切相关, 但在茶树上鲜见报道。本研究通过RACE和RT-PCR技术克隆获得茶树中的一个 bZIP基因, 它与其他植物bZIP具有较高的同源性, 序列分析表明含有bZIP转录因子家族典型的碱性结构域和亮氨酸拉链结构(氨基酸序列第95~127位), 属于B-zip1家族, 命名为 CsbZIP1。CsbZIP1蛋白与拟南芥不同亚家族bZIP蛋白的系统发育树分析显示其与F亚家族聚为一类。目前, 植物中关于F亚家族和B亚家族的研究报道较少, 但其他亚家族在ABA、逆境胁迫、种子发育、抗氧化、光信号转导、光合作用以及糖信号转导等方面发挥重要作用[ 4, 13, 24]; 磷酸化
位点预测结果表明CsbZIP1蛋白含有多个磷酸化位点, 可能与bZIP蛋白的活性调控有关。磷酸化和去磷酸化是ABA信号通路中的关键步骤, 低温、干旱和高盐等逆境胁迫能够刺激植物体内ABA含量增加, 从而激活SnRK2蛋白激酶, 将bZIP转录因子的保守域磷酸化, 从而使bZIP蛋白活化, 再通过与顺式作用元件结合调控基因表达, 以此来增强植物的抗逆性[ 3, 25, 26]。蛋白质三级结构预测显示, CsbZIP1蛋白具有高度保守的由α-螺旋和亮氨酸形成的bZIP结构域, 与报道的拟南芥AtbZIP蛋白的三级结构模型相似。
亚细胞定位预测显示CsbZIP1蛋白最可能定位于细胞核, 另外还可能定位在叶绿体、质体和胞液。为了进一步验证CsbZIP1定位在核内, 通过洋葱表皮亚细胞定位试验分析了它的亚细胞定位。试验结果与预测结果相近, CsbZIP1蛋白主要定位在细胞核, 同时在细胞的其他部位也能够检测到微弱信号。这与其他植物bZIP转录因子的亚细胞定位研究报道相一致[ 11, 14, 27]。
已有研究表明, bZIP转录因子在非生物胁迫, 如干旱、低温、高盐等中参与调控基因表达[ 2, 9, 13]。本研究用qRT-PCR分析 CsbZIP1在ABA、低温和盐胁迫下的表达模式。依据前人[ 19, 20, 21]报道的茶树胁迫处理结果及本实验室前期[ 22]的研究结果, 分别选用浓度100 μmol L-1和250 mmol L-1的ABA和NaCl处理茶树。低温是限制茶树生长的最重要影响因子之一, 但植物响应低温胁迫与ABA信号途径可能密切相关, 为了分析 CsbZIP1在低温和ABA处理后的表达模式变化, 我们选取在处理后0、2、8和24 h的样品试验。为了分析NaCl处理在短时内对表达的影响, 我们分析了处理后4 h的表达, 通过与0 h和24 h时的表达比较, 明确 CsbZIP1在短时(4 h)内的表达模式。结果表明, CsbZIP1对低温和ABA处理的响应模式不尽相同, ABA处理2 h表达被迅速抑制, 从2~24 h的过程中, 表达逐渐恢复到处理前的水平; 低温处理2 h则诱导其表达, 且在处理的24 h内表达均是上调的。说明在低温和ABA处理后, 短时内(2 h)调控作用相反, 但在2~24 h过程中能够诱导 CsbZIP1表达, 与处理前相比, 低温下的表达上调更显著, 表明 CsbZIP1在低温胁迫响应过程中起着重要作用。盐胁迫下, 4 h时的表达被显著上调, 24 h的表达比4 h低, 但仍比处理前0 h的表达量高, 推测在处理24 h内表达上调。Zou等[ 28]研究水稻 bZIP基因表明, 250 mmol L-1 NaCl处理24 h内的表达量是上调的, 且5 h的表达量最高。说明NaCl处理对 bZIP基因的诱导可能在4~5 h内就能够达到最大, 这为以后深入研究其在抗盐胁迫中的功能提供了参考。
大量的研究结果表明, 不同的 bZIP基因在逆境胁迫中的表达模式不尽相同。Liao等[ 8]将大豆 GmbZIP 44、 GmbZIP 46、 GmbZIP 62和 GmbZIP 784个基因转到拟南芥中过表达, 发现这4个基因均能增强对高盐和低温逆境的抗性, 而 GmbZIP 44、 GmbZIP 62和 GmbZIP 78的过表达能减弱ABA的敏感性, 表明这3个基因有可能通过上调 ABI1和 ABI2基因的表达来参与ABA信号途径, 对ABA、盐和低温逆境起负调控作用。Zou等[ 28]对水稻bZIP转录因子基因 OsABI5的表达研究表明, 4℃低温处理24 h后, 前5 h的表达量是下调的, 10 h后表达量逐渐上调。Orellana等[ 29]分离的番茄bZIP转录因子基因 SIAREB1同时受ABA和NaCl胁迫诱导, 100 μmol L-1 ABA处理24 h后, SIAREB1在叶片和根中的表达量是上升的; 300 mmol L-1 NaCl处理72 h后, 叶片和根中的表达量都是在6 h最大, 其后表达量降低。Ying等[ 11]克隆的玉米 ZmbZIP72基因的表达结果显示, NaCl胁迫(200 mmol L-1)处理24 h后, 表达量是增加的, 且在24 h表达量最大; 而4℃低温处理24 h后, 表达量变化不大。Gao等[ 6]研究大豆 GmbZIP1在不同逆境胁迫下的表达结果表明, 4℃低温处理24 h后, GmbZIP1的表达量是逐渐升高的, 24 h的表达量最大, 与本试验4℃低温表达模式相似。另外, Schlögl等[ 30]对甘蔗bZIP转录因子基因研究发现, 在100 μmol L-1 ABA处理12 h后, ScbZIP29和 ScbZIP31的表达量是上调的, 而 ScbZIP24的表达量是下调的, 这与本试验的ABA表达模式是相似的。可见, bZIP基因在ABA信号途径中可能起正调控作用, 也可能是负调控作用。 CsbZIP1可能还参与其他生物学功能, Cheng等[ 31]认为F亚家族的bZIP转录因子可能参与植物的光形态建成, 茶树bZIP的功能及其他家族基因有待后续发掘。但是, 从表达结果来看, CsbZIP1极有可能与茶树低温、盐、ABA胁迫等方面有密切关系。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] | |
| [25] | |
| [26] | |
| [27] | |
| [28] | |
| [29] | |
| [30] | |
| [31] |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}