关键词:玉米; 渗透胁迫; 基因表达谱; 差异表达基因 In silico Expression Profile of Maize Genes in Response to Osmotic Stress GE Shu-Juan1,**, SUN Ai-Qing2,**, LIU Peng2, ZHANG Jie-Dao1,*, DONG Shu-Ting2,* 1 State Key Laboratory of Crop Biology / Shandong Key Laboratory of Crop Biology / College of Life Sciences, Shandong Agricultural University, Tai’an 271018, China
2 College of Agriculture, Shandong Agricultural University, Tai’an 271018, China
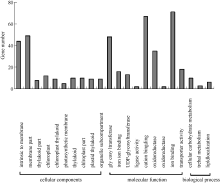
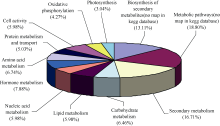
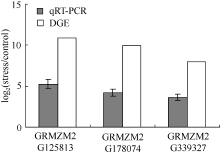
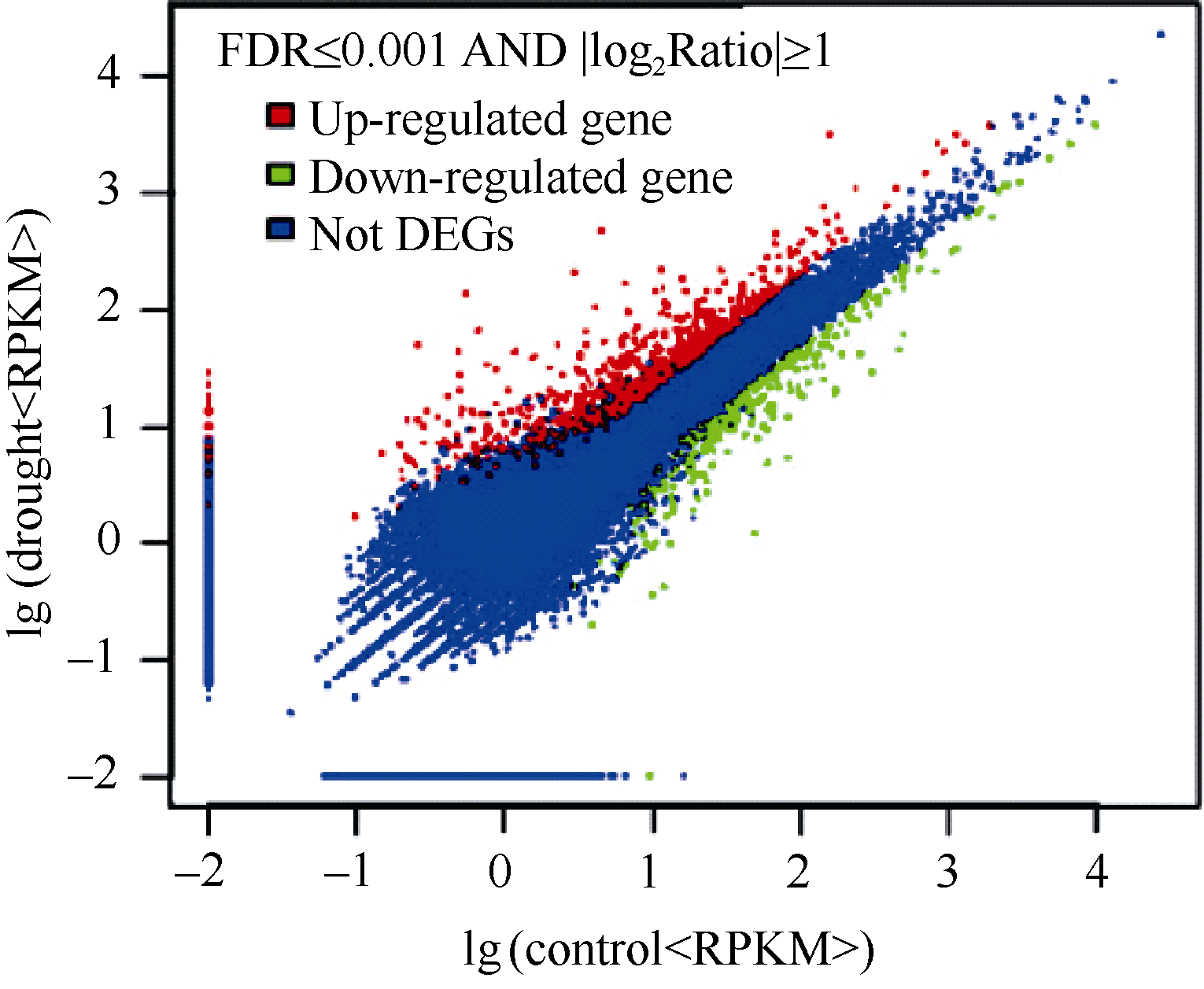
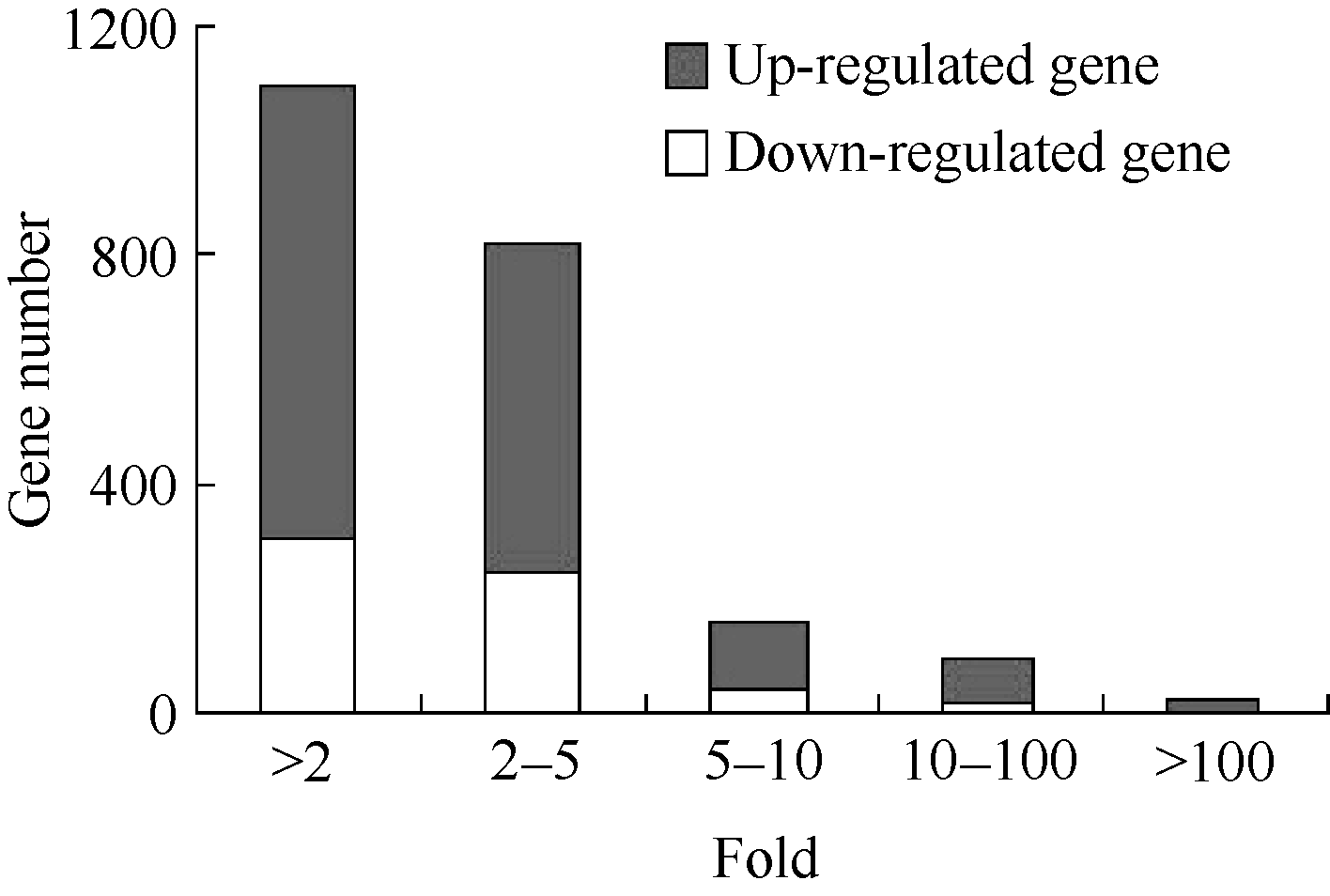
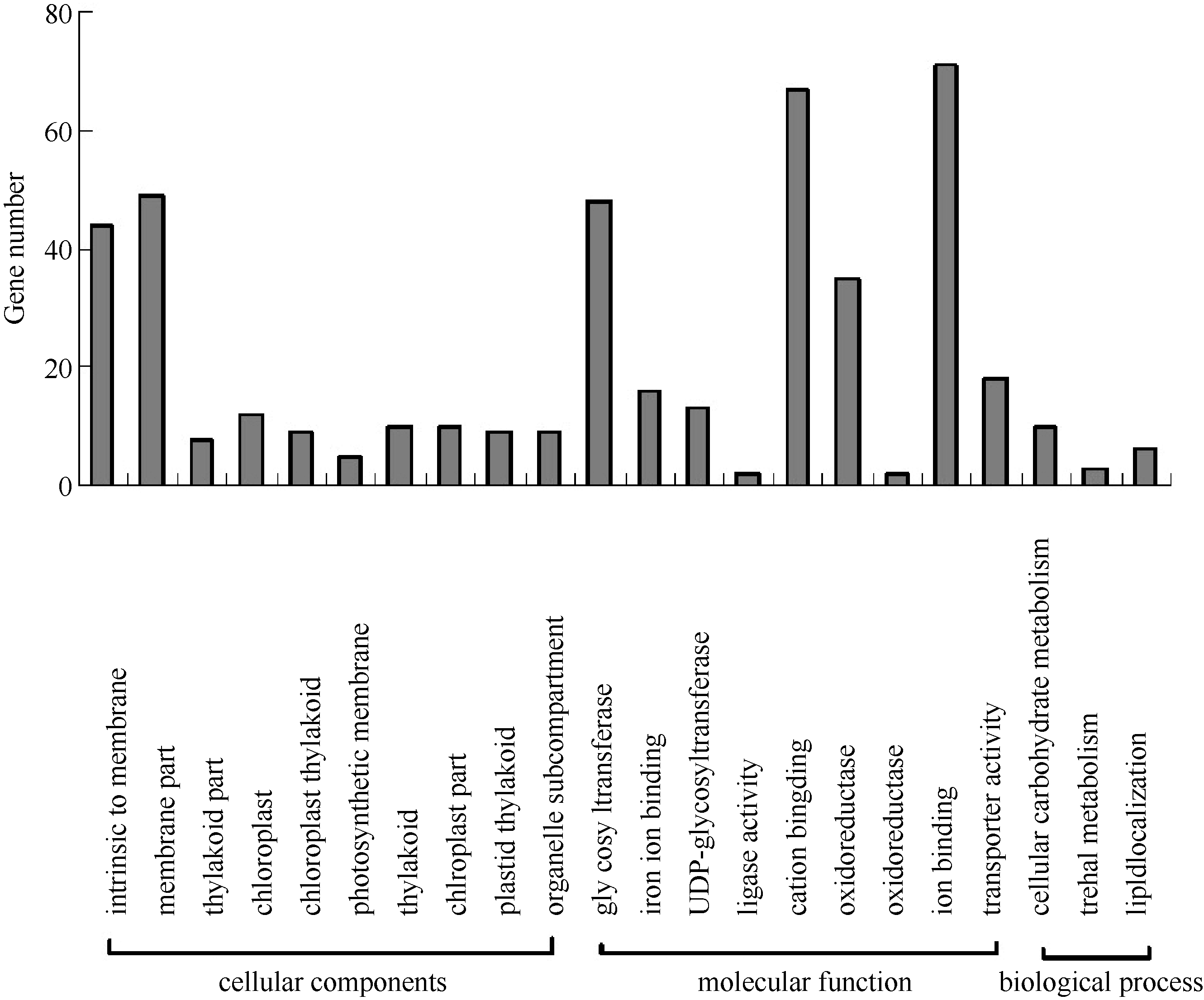
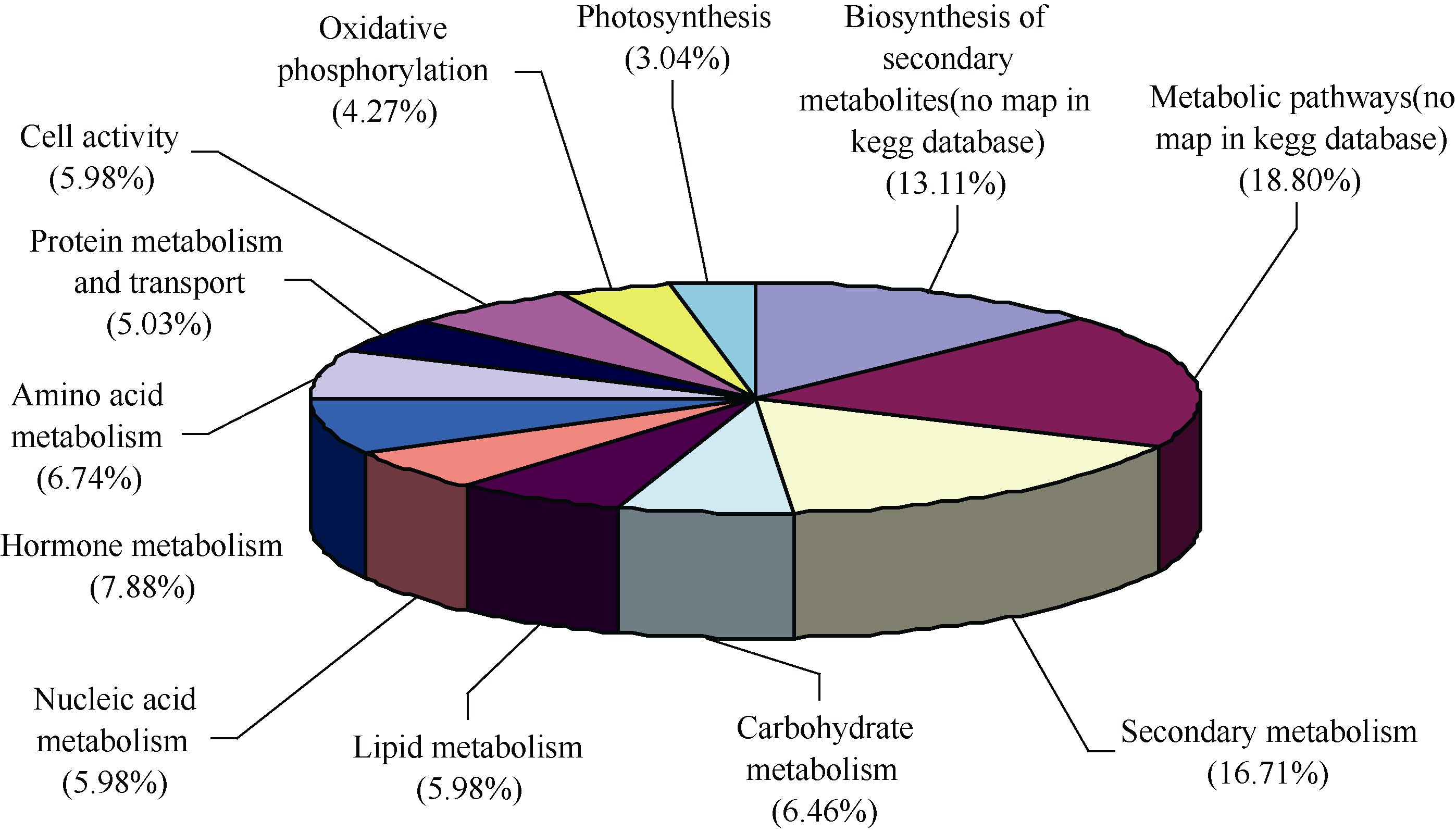
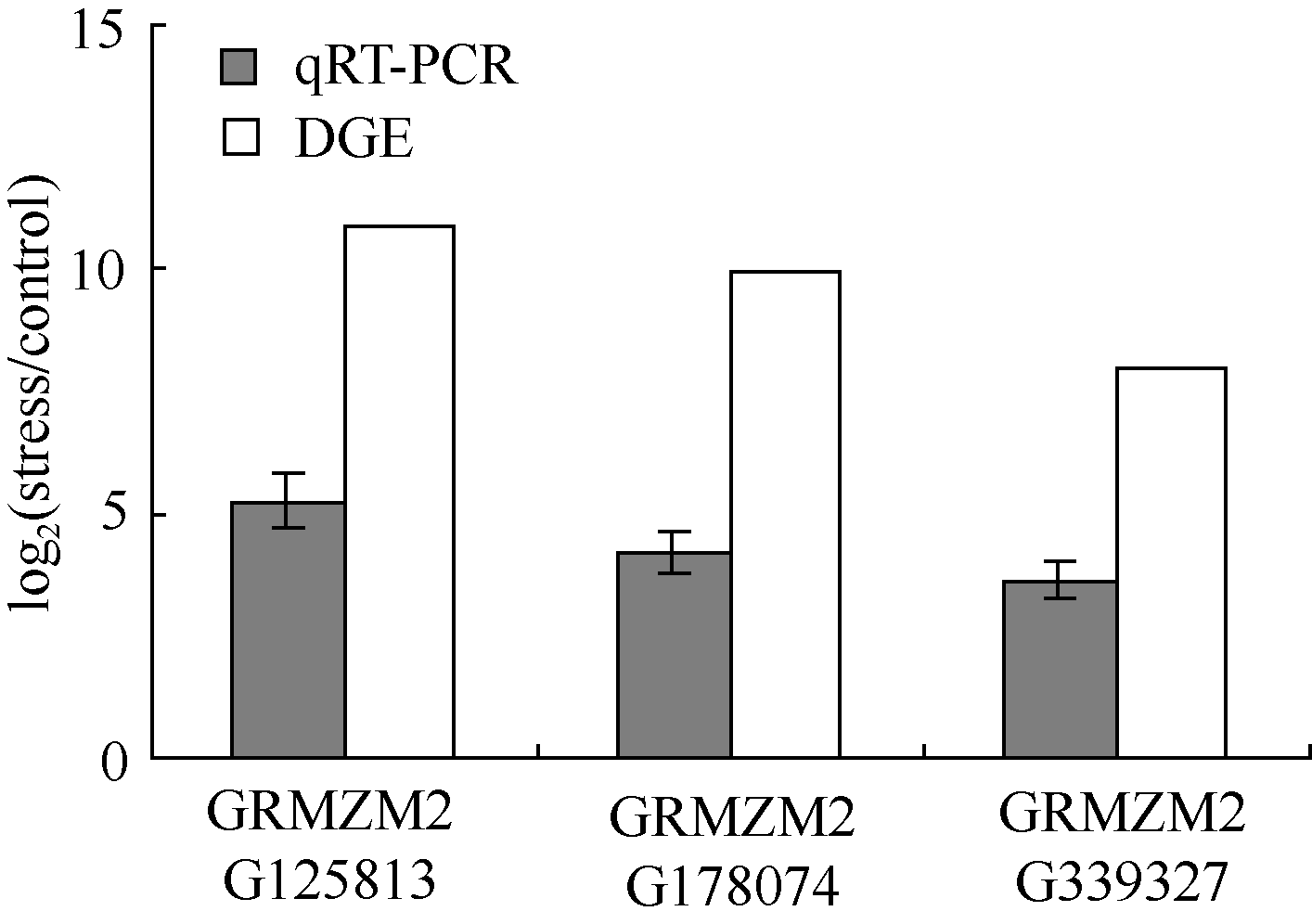
** Contributed equally to this work Fund: AbstractIn this study, a germplasm POB21 was used to analyze leaf cDNA library of maize treated with 15% PEG byin silico expression profile. The results indicated the POB21 genome shared significantly high similarity with reference genome. The gene expression in maize transcriptome presented strong nonhomogeneity and partial redundancy. A total of 1097 differentially expressed genes (DEGs) were screened out, of which 795 DEGs were up-regulated and 302 DEGs were down-regulated. GO enrichment analysis of these DEGs showed that three cellular components and three molecular functions of glycosyl transferase activity were enriched. The KEGG pathway analysis showed these DEGs were involved in metabolisms of carbohydrate, protein, nucleic acid, lipid, secondary metabolites, hormones and energy. The DEGs involving in proline metabolism indicated glutamate pathway is the predominant accumulation way of proline under osmotic stress in maize. The result lay a foundation for further study of molecular mechanism in response to osmotic response and functional genes screening of maize.
Keyword:Maize; Osmotic stress; Gene expression profile; Differentially expressed genes Show Figures Show Figures
张仁和, 薛吉全, 浦军, 赵兵, 张兴华, 郑友军, 卜令铎. 干旱胁迫对玉米苗期植株生长和光合特性的影响. 作物学报, 2011, 37: 521-528ZhangR H, XueJ Q, PuJ, ZhaoB, ZhangX H, ZhengY J, BuL D. Influence of drought stress on plant growth and photosynthetic traits in maize seedlings. Acta Agron Sin, 2011, 37: 521-528 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[2]
MunnsR, TesterM. Mechanisms of salinity tolerance. Annu Rev Plant Biol, 2008, 59: 651-681[本文引用:1][JCR: 25.962]
[3]
刘志玲, 程丹. 植物抗旱生理研究进展与育种. 中国农学通报, 2011, 27(24): 249-252LiuZ L, ChengD. Plant drought-resistant physiology research progress and breeding. Chin Agric Sci Bull, 2011, 27(24): 249-252 (in Chinese with English abstract)[本文引用:1]
[4]
张红, 董树亭. 玉米对盐胁迫的生理响应及抗盐策略研究进展. 玉米科学, 2011, 19(1): 64-69ZhangH, DongS T. Research progress on the physiological and biochemistry responses of salt tolerance and strategies of salt resistance in maize. J Maize Sci, 2011, 19(1): 64-69 (in Chinese with English abstract)[本文引用:1][CJCR: 0.965]
[5]
KongX, SunL, ZhouY, ZhangM, LiuY, PanJ, LiD. ZmMKK4 regulates osmotic stress through reactive oxygen species scavenging in transgenic tobacco. Plant Cell Rep, 2011, 30: 2097-3104[本文引用:1][JCR: 2.509]
[6]
YingS, ZhangD F, FuJ, ShiY S, SongY C, WangT Y, LiY. Cloning and characterization of a maize bZIP transcription factor, ZmbZIP72, confers drought and salt tolerance in transgenic Arabidopsis. Planta, 2012, 235: 253-266[本文引用:1][JCR: 3.347]
[7]
LiuY, WangL, XingX, SunL, PanJ, KongX, ZhangM, LiD. ZmLEA3, a multifunctional group 3 LEA protein from maize (Zea mays L. ), is involved in biotic and abiotic stresses. Plant Cell Physiol, 2013, 54: 944-959[本文引用:1][JCR: 4.134]
[8]
李会勇, 黄素华, 赵久然, 王凤格, 张中保, 毛毅辉, 王秀堂, 石云素, 宋燕春, 王国英, 黎裕, 王天宇. 应用抑制差减杂交法分离玉米幼苗期叶片土壤干旱诱导的基因. 中国农业科学, 2007, 40: 882-888LiH Y, HuangS H, ZhaoJ R, WangF G, ZhangZ B, MaoY H, WangX T, ShiY S, SongY C, WangG Y, LiY, WangT Y. Isolating soil drought-induced genes from maize seedlings leaves through suppression subtractive hybridization. Sci Agric Sin, 2007, 40: 882-888 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[9]
滕晓坤, 肖华胜. 基因芯片与高通量DNA测序技术前景分析. 中国科学: C辑, 2008, 38: 891-899TengX Q, XiaoH S. The prospect analysis of gene chip and high-throughput DNA sequencing. Sci Sin Vitae, 2008, 38: 891-899 (in Chinese)[本文引用:1][CJCR: 0.833]
KanehisaM, ArakiM, GotoS, HattoriM, HirakawaM, ItohM, KatayamaT, KawashimaS, OkudaS, TokimatsuT, YamanishiY. KEGG for linking genomes to life and the environment. Nucl Acids Res, 2008, 36: 480-484[本文引用:1]
[12]
LiH W, ZangB S, DengX W, WangX P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta, 2011, 234: 1007-1018[本文引用:1][JCR: 3.347]
[13]
NishizawaA, YabutaY, ShigeokaS. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol, 2008, 147: 1251-1263[本文引用:1][JCR: 6.555]
[14]
AmiardV, Morvan-Bertrand A, BillardJ P, HuaultC, KellerF, Prud’hommeM P. Fructans, but not the sucrosyl-galactosides, raffinose and loliose, are affected by drought stress in perennial ryegrass. Plant Physiol, 2003, 132: 2218-2229[本文引用:1][JCR: 6.555]
[15]
MonrealJ A, Arias-BaldrichC, Pérez-MontañoF, Gand ulloJ, EchevarríaC, García-MauriñoS. Factors involved in the rise of phosphoenolpyruvate carboxylase-kinase activity caused by salinity in sorghum leaves. Planta, 2013, 237: 1401-1413[本文引用:1][JCR: 3.347]
[16]
刘芳, 卢长明. 植物非特异脂质转运蛋白研究现状与展望. 遗传, 2013, 35: 307-314LiuF, LuC M. An overview of non-specific lipid transfer protein in plant. Hereditas (Beijing), 2013, 35: 307-314 (in Chinese with English abstract)[本文引用:1]
[17]
GuoC, GeX, MaH. The rice OsDIL gene plays a role in drought tolerance at vegetative and reproductive stages. Plant Mol Biol, 2013, 82: 239-253[本文引用:1][JCR: 3.518]
[18]
HuangY, LemieuxM J, SongJ, AuerM, WangD N. Structure and mechanism of the glycerol-3-phosphate transporter from Escherichia coli. Science, 2003, 301: 616-620[本文引用:1]
[19]
LanP, LiW, WangH, MaW. Characterization, sub-cellular localization and expression profiling of the isoprenylcysteine methylesterase gene family in Arabidopsis thaliana. BMC Plant Biol, 2010, 10: 212[本文引用:1][JCR: 4.354]
[20]
KoopsP, PelserS, IgnatzM, KloseC, Marrocco-SeldenK, KretschT. EDL3 is an F-box protein involved in the regulation of abscisic acid signalling in Arabidopsis thaliana. J Exp Bot, 2011, 62: 5547-5560[本文引用:1][JCR: 5.242]
LuM, YingS, ZhangD F, ShiY S, SongY C, WangT Y, LiY. A maize stress-responsive NAC transcription factor, ZmSNAC1, confers enhanced tolerance to dehydration in transgenic Arabidopsis. Plant Cell Rep, 2012, 31: 1701-1711[本文引用:1][JCR: 2.509]
[23]
DietzK J, VogelM O, ViehhauserA. AP2/EREBP transcription factors are part of gene regulatory networks and integrate metabolic, hormonal and environmental signals in stress acclimation and retrograde signalling. Protoplasma, 2010, 245: 3-14[本文引用:1][JCR: 2.855]
[24]
GuoZ J, ChenX J, WuX L, LingJ Q, XuP. Overexpression of the AP2/EREBP transcription factor OPBP1 enhances disease resistance and salt tolerance in tobacco. Plant Mol Biol, 2004, 55: 607-618[本文引用:1][JCR: 3.518]
[25]
GaoS Q, ChenM, XiaL Q, XiuH J, XuZ S, LiLC, ZhaoC P, ChengX G, MaY Z. A cotton (Gossypium hirsutum) DRE-binding transcription factor gene, GhDREB, confers enhanced tolerance to drought, high salt, and freezing stresses in transgenic wheat. Plant Cell Rep, 2009, 28: 301-311[本文引用:1][JCR: 2.509]
[26]
ZhaoY, ZhouY, JiangH, LiX, GanD, PengX, ZhuS, ChengB. Systematic analysis of sequences and expression patterns of drought-responsive members of the HD-Zip gene family in maize. PLoS One, 2011, 6(12): e28488: 1-14[本文引用:1][JCR: 3.73]
[27]
CabelloJ V, ChanR L. The homologous homeodomain-leucine zipper transcription factors HaHB1 and AtHB13 confer tolerance to drought and salinity stresses via the induction of proteins that stabilize membranes. Plant Biotechnol J, 2012, 10: 815-825[本文引用:1][JCR: 6.279]
[28]
LiY S, SunH, WangZ F, DuanM, HuangS D, YangJ, HuangJ, ZhangH S. A novel nuclear protein phosphatase 2C negatively regulated by ABL1 is involved in abiotic stress and panicle development in rice. Mol Biotechnol, 2013, 54: 703-710[本文引用:1][JCR: 2.262]
[29]
LiuX, ZhuY, ZhaiH, CaiH, JiW, LuoX, LiJ, BaiX. AtPP2CG1, a protein phosphatase 2C, positively regulates salt tolerance of Arabidopsis in abscisic acid-dependent manner. Biochem Biophys Res Commun, 2012, 422: 710-715[本文引用:1][JCR: 2.406]
[30]
HuX, LiuL, XiaoB, LiD, XingX, KongX, LiD. Enhanced tolerance to low temperature in tobacco by over-expression of a new maize protein phosphatase 2C, ZmPP2C2. J Plant Physiol, 2010, 167: 1307-1315[本文引用:1][JCR: 2.699]
[31]
WatanabeN, LamE. Arabidopsis Bax inhibitor-1 functions as an attenuator of biotic and abiotic types of cell death. Plant J, 2006, 45: 884-894[本文引用:1][JCR: 6.582]
[32]
KimD Y, JinJ Y, Alejand roS, MartinoiaE, LeeY. Overexpression of AtABCG36 improves drought and salt stress resistance in Arabidopsis. Physiol Plant, 2010, 139: 170-180[本文引用:1][JCR: 6.555]
[33]
PangK, LiY, LiuM, MengZ, YuY. Inventory and general analysis of the ATP-binding cassette (ABC) gene superfamily in maize (Zea mays L. ). Gene, 2013, 526: 411-428[本文引用:1][JCR: 2.196]
[34]
WilkinsonS, KudoyarovaG R, VeselovD S, ArkhipovaT N, DaviesW J. Plant hormone interactions: innovative targets for crop breeding and management. J Exp Bot, 2012, 63: 3499-3509[本文引用:1][JCR: 5.242]
RizhskyL, LiangH, ShumanJ, ShulaevV, DavletovaS, MittlerR. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol, 2004, 134: 1683-1696[本文引用:1][JCR: 6.555]
[37]
AhmadP, JaleelC A, SalemM A, NabiG, SharmaS. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit Rev Biotechnol, 2010, 30: 161-175[本文引用:1][JCR: 5.095]
[38]
HouryW A. Chaperone-assisted protein folding in the cell cytoplasm. Curr Protein Pept Sci, 2001, 2: 227-244[本文引用:1][JCR: 2.326]
 , 董树亭
, 董树亭


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}