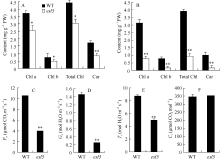
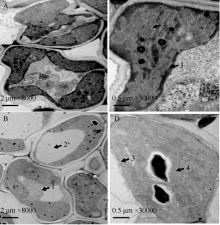
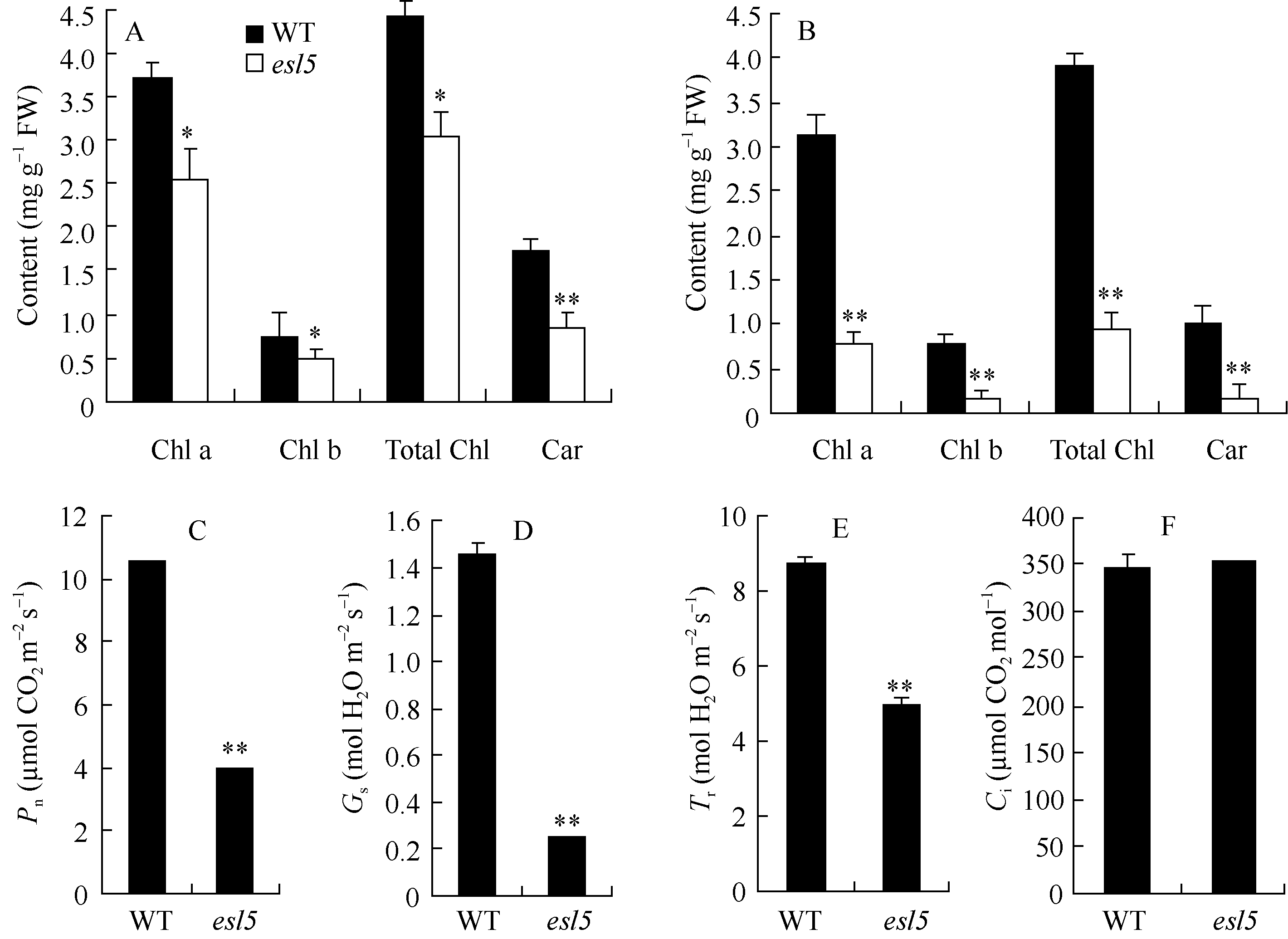
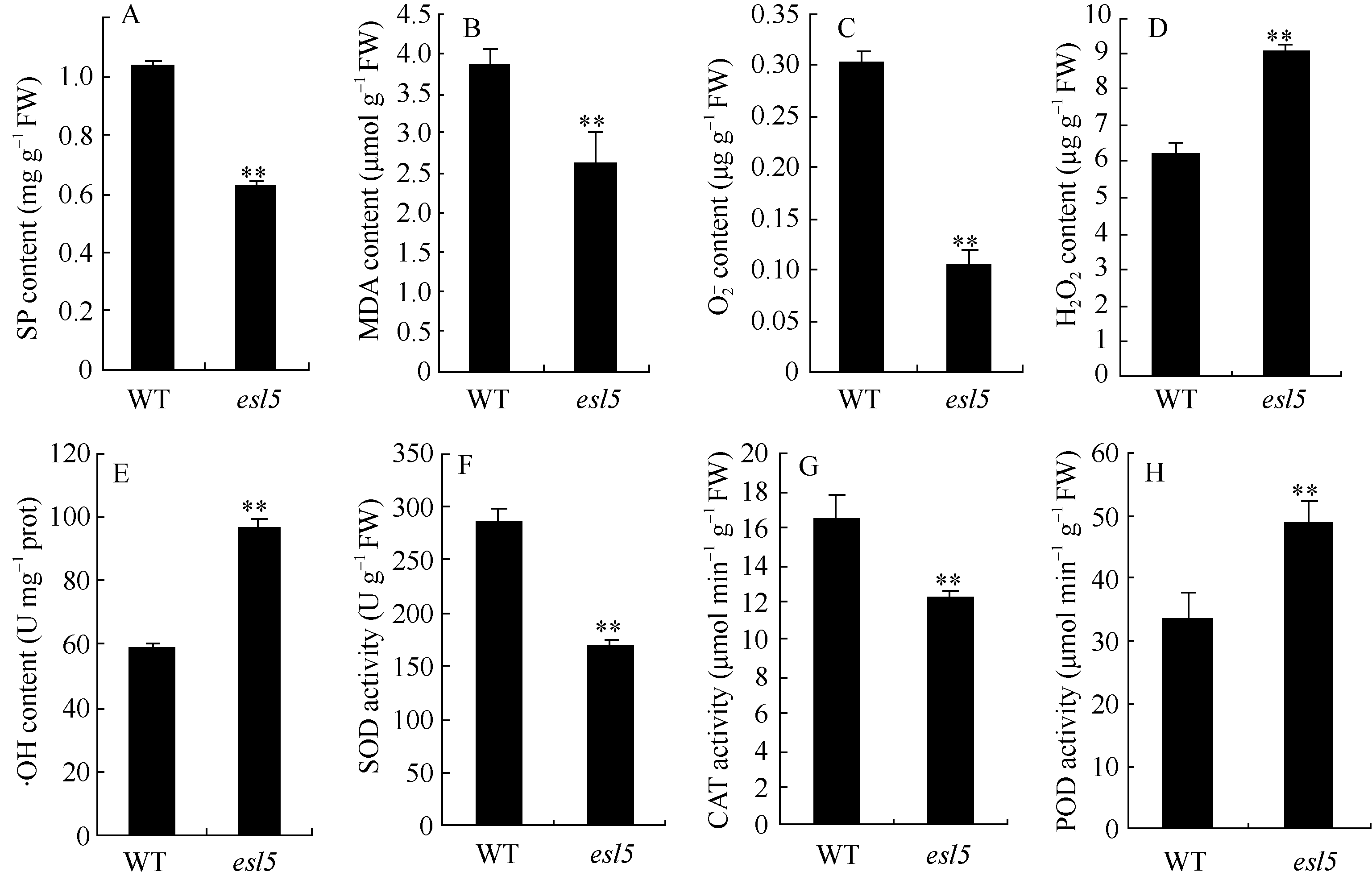
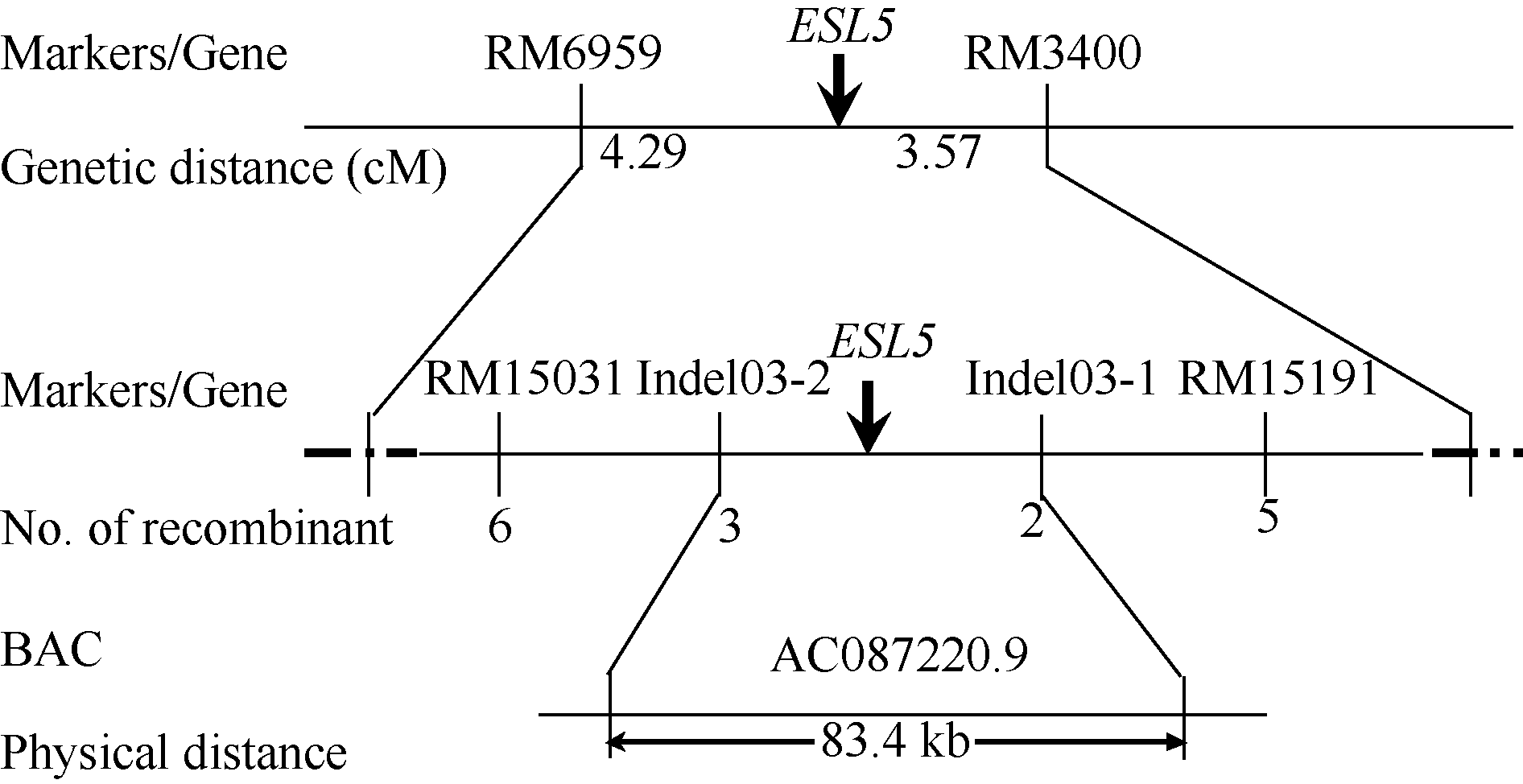
关键词:水稻(Oryza sativa); 早衰; 基因定位; 活性氧(ROS); 光系统 Identification and Gene Fine Mapping of Early Senescent Leaf Mutantesl5 inOryza sativa SANG Xian-Chun, XU Fang-Fang, ZHU Xiao-Yan, XING Ya-Di, HE Pei-Long, ZHANG Chang-Wei, YANG Zheng-Lin, HE Guang-Hua* Rice Research Institute of Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops /Engineering Research Center of South Upland Agriculture, Ministry of Education, Chongqing 400716, China Fund: AbstractPremature senility directly influences crop yield and quality in the production. Therefore, it is crucial to identify early senescent leaf mutants and then clone genes associated with senescence by map-based strategy, which is significant in the research of senescence mechanism. An early senescent leaf mutantesl5 has been discovered from the progeny ofindica restorer line Jinhui10 with seeds treated by ethylmethane sulfonate, this paper performed the studies on its morphological identification, cytological observation, physiological analysis and gene mapping. The results showed that theesl5 demonstrated normal phenotype at the seedling stage, displayed yellow green color at the tillering stage, and appeared yellow and senescent leaf blades at booting stage. Cell structure of theesl5 was badly influenced, showing destroyed cell membranes, loose and irregular stroma lamella in the section of senescent leaves. Chlorophyll content and photosynthetic rate of theesl5 were significantly lower than those of the wild type. Besides of the lower activities of SOD and CAT, the higher contents of ·OH and H2O2 were detected in theesl5 than in the wild type. The growth period ofesl5 delayed 20 days, grain number per panicle, filled-seed number and seed setting rate decreased significantly, but 1000-seed weight increased significantly at compared with the wild type. Genetic analysis suggested that the mutant trait was controlled by a recessive nuclear gene, and theESL5 was fine mapped on chromosome 3 between Indel markers Indel03-1 and Indel03-2 with 83.4 kb physical distance, containing 11 annotated genes. These results provide a foundation for the cloning ofESL5 by map-based strategy, which is essential in the research of senescence mechanism as well as molecular breeding inOryza sativa.
Keyword:Rice (Oryza sativa); Early senescence; Gene mapping; Reactive oxygen species (ROS); Photosynthetic systems Show Figures Show Figures
AnsariM I, ChenS C G. Leaf senescence: an overview. Intl J Comput Sci Telecommun, 2011, 1: 110-114[本文引用:1]
[2]
LimP O, KimH J, NamH G. Leaf senescence. Annu Rev Plant Biol, 2007, 58: 115-136[本文引用:1][JCR: 25.962]
[3]
WuX Y, KuaiB K, JiaJ Z, JingH C. Regulation of leaf senescence and crop genetic improvement. J Integr Plant Biol, 2012, 54: 936-952[本文引用:1][JCR: 3.75][CJCR: 0.7555]
[4]
KongZ, LiM, YangW, XuW, XueY. A novel nuclear-localized CCCH-type zinc finger protein, OsDOS, is involved in delaying leaf senescence in rice. Plant Physiol, 2006, 141: 1376-1388[本文引用:1][JCR: 6.555]
[5]
GuoY F. Towards systems biological understand ing of leaf senescence. Plant Mol Biol, 2013, 82: 519-528[本文引用:1][JCR: 3.518]
[6]
Buchanan-WollastonV, PageT, HarrisonE, BreezeE, LimP O, NamH G, LinJ F, WuS H, SwidzinskiJ, IshizakiK, LeaverC J. Comparative transcriptome analysis reveals significant differences in gene expression and signaling pathways between developmental and dark/starvation-induced senescence in Arabidopsis. Plant J, 2005, 42: 567-585[本文引用:1][JCR: 6.582]
[7]
LiuL, ZhouY, ZhouG, YeR, ZhaoL, LiX, LinY. Identification of early senescence-associated genes in rice flag leaves. Plant Mol Biol, 2008, 67: 37-55[本文引用:1][JCR: 3.518]
[8]
WangN L, LongT A, YaoW, XiongL Z, ZhangQ F, WuC Y. Mutant resources for the functional analysis of the rice genome. Mol Plant, 2013, 6: 596-604[本文引用:1][JCR: 6.126][CJCR: 0.9487]
[9]
WangJ, WuS J, ZhouY, ZhouL H, XuJ F, HuJ, FangY X, GuM H, LiangG H. Genetic analysis and molecular mapping of a presenescing leaf gene psl1 in rice (Oryza sativa L. ). Chin Sci Bull, 2006, 51: 2986-2992[本文引用:2][CJCR: 0.95]
[10]
ZhuL, LiuW Z, WuC, LuanW J, FuY P, HuG C, SiH M, SunZ X. Identification and fine mapping of a gene related to pale green leaf phenotype near centromere region in rice (Oryza sativa L. ). Rice Sci, 2007, 14: 172-180[本文引用:2][CJCR: 0.3579]
[11]
FangL K, LiY F, GongX P, SangX C, LingY H, WangX W, CongY F, HeG H. Genetic analysis and gene mapping of dominant presenescing leaf gene PSL3 in rice (Oryza sativa L. ). Chin Sci Bull, 2010, 55: 2517-2521[本文引用:2][CJCR: 0.95]
[12]
LiF Z, HuG C, FuY P, SiH M, BaiX M, SunZ X. Genetic analysis and high-resolution mapping of a premature senescence gene Pse(t) in rice (Oryza sativa L. ). Genome, 2005, 48: 738-746[本文引用:2][JCR: 1.668]
[13]
YangY L, RaoY C, LiuH J, FangY X, DongG J, HuangL C, LengY J, GuoL B, ZhangG H, HuJ, GaoZ Y, QianQ, ZengD L. Characterization and fine mapping of an early senescence mutant (es-t) in Oryza sativa L. Chin Sci Bull, 2011, 56: 2437-2443[本文引用:2][CJCR: 0.95]
[14]
徐芳芳, 桑贤春, 任德勇, 唐彦强, 胡宏伟, 杨正林, 赵芳明, 何光华. 水稻早衰突变体esl2的遗传分析和基因定位. 作物学报, 2012, 38: 1347-1353XuF F, SangX C, RenD Y, TangY Q, HuH W, YangZ L, ZhaoF M, HeG H. Genetic analysis and gene mapping of early senescence leaf mutant esl2 in rice. Acta Agron Sin, 2012, 38: 1347-1353 (in Chinese with English abstract)[本文引用:3][CJCR: 1.667]
[15]
苗润隆, 蒋钰东, 廖红香, 徐芳芳, 何光华, 杨正林, 赵芳明, 桑贤春. 水稻早衰突变体esl3的鉴定与基因定位. 作物学报, 2013, 39: 862-867MiaoR L, JiangY D, LiaoH X, XuF F, HeG H, YangZ L, ZhaoF M, SangX C. Identification and gene mapping of rice early senescent leaf (esl3) mutant. Acta Agron Sin, 2013, 39: 862-867 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[16]
杜青, 方立魁, 桑贤春, 凌英华, 李云峰, 杨正林, 何光华, 赵芳明. 水稻叶尖早衰突变体lad的形态、生理分析与基因定位. 作物学报, 2012, 38: 168-173DuQ, FangL K, SangX C, LingY H, LiY F, YangZ L, HeG H, ZhaoF M. Analysis of phenotype and physiology of leaf apex dead mutant (lad) in rice and mapping of mutant gene. Acta Agron Sin, 2012, 38: 168-173 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[17]
YanW Y, YeS H, JinQ S, ZengL J, PengY, YanD W, YangW B, YangD L, HeZ H, DongY J, ZhangX M. Characterization and mapping of a novel mutant sms1 (senescence and male sterility 1) in rice. J Genet Genomics, 2010, 37: 47-55[本文引用:2][JCR: 2.076][CJCR: 1.323]
[18]
LinA H, WangY Q, TangJ Y, XueP, LiC L, LiuL C, HuB, YangF Q, LoakeG, ChuC C. Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide induced leaf cell death in rice. Plant Physiol, 2012, 158: 1451-1464[本文引用:1][JCR: 6.555]
[19]
ShiB, NiL, ZhangA Y, CaoJ M, ZhangH, QinT T, TanM P, ZhangJ H, JiangM Y. OsDMI3 is a novel component of abscisic acid signaling in the induction of antioxidant defense in leaves of rice. Mol Plant, 2012, 5: 1359-1374[本文引用:1][JCR: 6.126][CJCR: 0.9487]
[20]
JiaoB B, WangJ J, ZhuX D, ZengL J, LiQ, HeZ H. A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice. Mol Plant, 2012, 5: 205-217[本文引用:1][JCR: 6.126][CJCR: 0.9487]
[21]
QiaoY, JiangW, LeeJ, ParkB, ChoiM S, PiaoR, WooM O, RohJ H, HanL Z, PaekN C, SeoH S, KohH J. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit mu1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol, 2010, 185: 258-274[本文引用:1][JCR: 6.736]
[22]
WellburnA R. The spectral determination of chlorophyll a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J Plant Physiol, 1994, 144: 307-313[本文引用:1][JCR: 2.699]
[23]
桑贤春, 何光华, 张毅, 杨正林, 裴炎. 水稻PCR扩增模板的快速制备. 遗传, 2003, 25: 705-707SangX C, HeG H, ZhangY, YangZ L, PeiY. The simple gain of templates of rice genomes DNA for PCR. Hereditas (Beijing), 2003, 25: 705-707 (in Chinese with English abstract)[本文引用:1]
[24]
SedighehH G, MortazavianM, NorouzianD, AtyabiM, AkbarzadehA, HasanpoorK, GhorbaniM. Oxidative stress and leaf senescence. BMC Res Notes, 2011, 4: 477[本文引用:1]
[25]
RayP D, HuangB W, TsujiY. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal, 2012, 24: 981-990[本文引用:1][JCR: 4.304]
[26]
D’AutréauxB, ToledanoM B. ROS as signalling molecules: mechanisms that generate specificity in ROS homeostasis. Mol Cell Biol, 2007, 8: 813-824[本文引用:1][JCR: 5.372]
[27]
MorimotoH, IwataK, OgonukiN, InoueK, AtsuoO, Kanatsu-Shinohara M, MorimotoT, Yabe-NishimuraC, ShinoharaT. ROS are required for mouse spermatogonial stem cell self-renewal. Cell Stem Cell, 2013, 12: 774-786[本文引用:1][JCR: 25.315]
[28]
LiuJ, FinkeT. Stem cells and oxidants: too little of a bad thing. Cell Metab, 2013, 18: 1-2[本文引用:1][JCR: 14.619]
HungK T, KaoC H. Hydrogen peroxide is necessary for abscisic acid-induced senescence of rice leaves. J Plant Physiol, 2004, 161: 1347-1357[本文引用:1][JCR: 2.699]
[31]
FuchsS, GrillE, MeskieneI, SchweighoferA. Type 2C protein phosphatases in plants. FEBS J, 2013, 280: 681-693[本文引用:1][JCR: 4.25]
[32]
GuoY, GanS. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J, 2006, 46: 601-612[本文引用:1][JCR: 6.582]
[33]
ZhangK, GanS S. An abscisic acid-AtNAP transcription factor-SAG113 protein phosphatase 2C regulatory chain for controlling dehydration in senescing Arabidopsis leaves. Plant Physiol, 2012, 158: 961-969[本文引用:1][JCR: 6.555]
[34]
ZhangK, XiaX, ZhangY, GanS S. An ABA-regulated and golgi-localized protein phosphatase controls water loss during leaf senescence in Arabidopsis. Plant J, 2012, 69: 667-678[本文引用:1][JCR: 6.582]
[35]
秘彩莉, 刘旭, 张学勇. F-box蛋白质在植物生长发育中的功能. 遗传, 2006, 28: 1337-1342BeiC L, LiuX, ZhangX Y. The function of f-box protein in plant growth and development. Hereditas (Beijing), 2006, 28: 1337-1342 (in Chinese with English abstract)[本文引用:1]
[36]
WooH R, ChungK M, ParkJ H, OhS A, AhnT, HongS H, JangS K, NamH G. ORE9, an F-Box protein that regulates leaf senescence in Arabidopsis. Plant Cell, 2001, 13: 1779-1790[本文引用:1][JCR: 9.251]
 , 徐芳芳, 朱小燕, 邢亚迪, 何沛龙, 张长伟, 杨正林, 何光华
, 徐芳芳, 朱小燕, 邢亚迪, 何沛龙, 张长伟, 杨正林, 何光华





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}