关键词:SSR; 蚕豆; 遗传多样性; 核心种质; 微核心种质 Genetic Diversity Analysis of Germplasm Resources and Construction of Mini-core Collections forVicia faba L. at Home and Abroad JIANG Jun-Ye1, YANG Tao1, WANG Fang1, FANG Li1, ZHONG Wei-Wen2, GUAN Jian-Ping1, ZONG Xu-Xiao1,* 1National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
2College of Life Science and Technology, Gansu Agricultural University, Lanzhou 730070, China
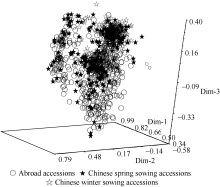
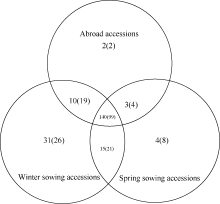
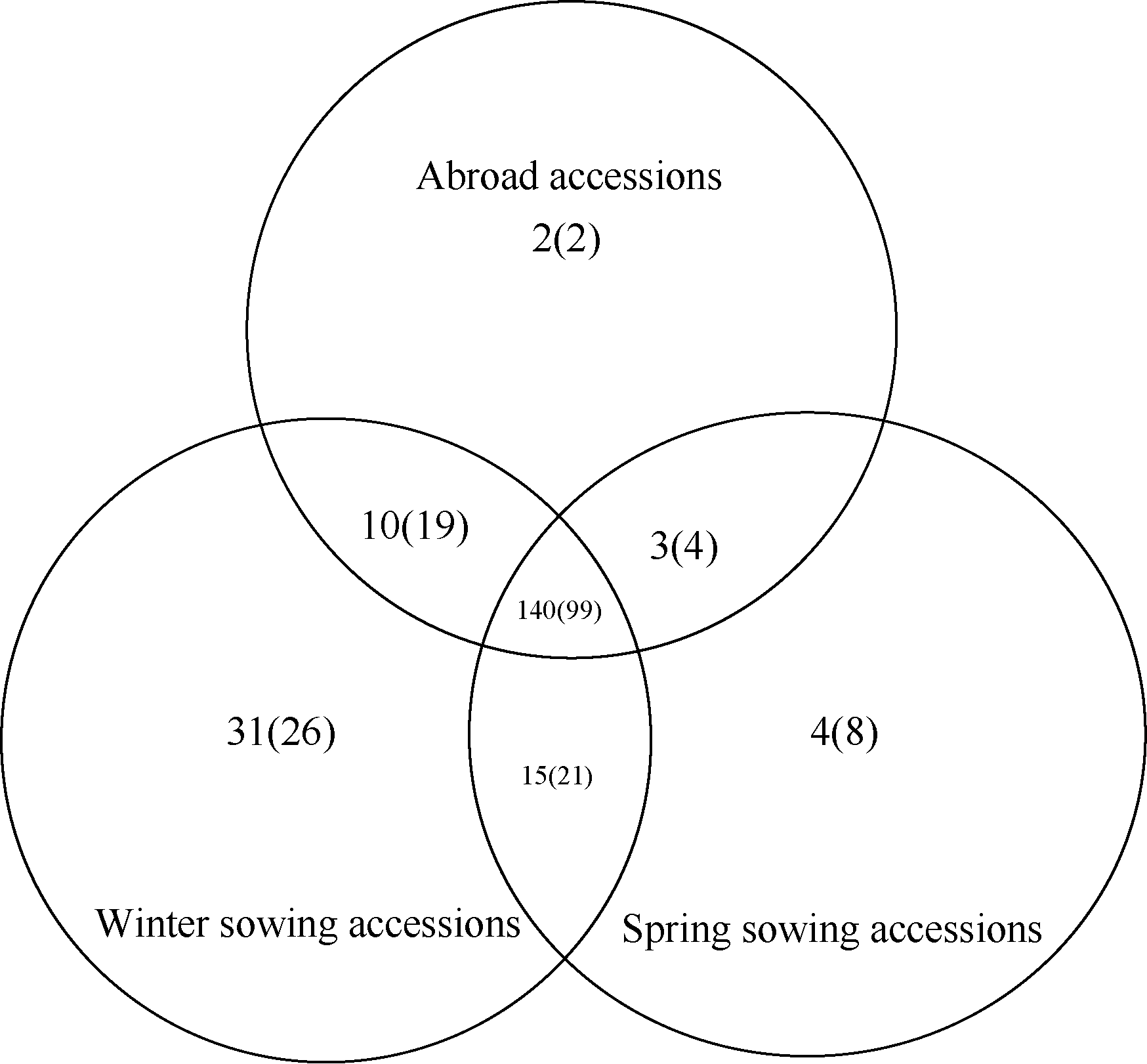
Fund: AbstractThe genetic diversity of 1075 genotypes from a primary geographic core collection of faba bean (Vicia faba L.) was analyzed by using SSR markers. The number of observed and effective alleles of 8.54 and 2.26, the Shannon’s information index of 1.02, were detected. Unconspicuous population structure among the 1075 genotypes indicated an extensive genetic background and nice representativeness. A mini-core collection comprised 129 Chinese and 63 oversea faba bean genotypes was randomly sampled from each cluster of dendrogram including all the 1075 tested genotypes. The observed alleles, effective alleles and Shannon’s information index of the mini-core collection covered 87.32%, 101.26%, and 101.82% of the primary geographic core collection respectively. The t-test did not show significant differences in genetic diversity between the primary geographic core collection and the mini-core collection, which validated the representativeness of the mini-core collection in genetic diversity of faba bean. The mini-core collection will play an important role in exploration of new genes within faba bean.
Keyword:SSR; Faba bean; Genetic diversity; Geographic core collection; Mini-core collection Show Figures Show Figures
表1 24对SSR引物在全部参试资源以及微核心种质中扩增的等位变异数、有效变异数和Shannon’s信息指数 Table 1 Alleles, effective alleles, and polymorphic information of SSR amplified products in all tested genotypes and mini-corecollections of faba bean
SSR位点 SSR primer pair
等位变异数 Number of alleles ( Na)
有效等位变异数 Number of effective alleles ( Ne)
有效等位变异所占比重 Ratio of Ne/Na
Shannon’s信息指数 Shannon’s information index ( I)
All accessions
Mini-core
All accessions
Mini-core
All accessions
Mini-core
All accessions
Mini-core
SSR6221
8
7
1.91
2.03
0.24
0.29
1.03
1.07
SSR5594
5
4
2.16
2.11
0.43
0.53
0.90
0.89
SSR2577
6
5
2.42
2.40
0.40
0.48
1.03
1.01
SSR6613
2
2
1.83
1.71
0.91
0.85
0.64
0.60
SSR1929
10
9
2.85
2.98
0.28
0.33
1.43
1.47
SSR3637
6
6
1.26
1.30
0.21
0.22
0.49
0.55
SSR5078
6
6
2.46
2.34
0.41
0.39
1.08
1.01
SSR2992
5
5
2.08
2.09
0.41
0.42
0.90
0.92
SSR9810
9
7
3.28
3.24
0.36
0.46
1.42
1.44
SSR10581
25
20
5.01
4.93
0.20
0.25
1.97
1.95
SSR4056
9
9
2.71
2.86
0.30
0.32
1.32
1.37
SSR10177
9
8
1.73
1.76
0.19
0.22
0.89
0.90
SSR3372
15
11
4.36
4.25
0.29
0.39
1.77
1.75
SSR1788
9
8
1.94
2.00
0.22
0.25
1.07
1.07
SSR5740
9
7
1.83
2.03
0.20
0.29
0.90
1.00
SSR4798
16
14
2.47
2.61
0.15
0.19
1.37
1.46
SSR3581
9
9
1.68
1.76
0.19
0.20
0.91
1.00
SSR2364
9
8
1.40
1.45
0.16
0.18
0.68
0.73
SSR6092
4
4
1.21
1.17
0.30
0.29
0.37
0.33
SSR4076
4
2
1.86
1.85
0.47
0.93
0.68
0.65
SSR6116
4
4
1.54
1.57
0.39
0.39
0.59
0.61
SSR102
9
9
2.96
3.17
0.33
0.35
1.37
1.44
SSR10296
6
5
1.82
1.78
0.30
0.36
0.91
0.89
SSR10421
11
10
1.38
1.41
0.13
0.14
0.68
0.73
合计 Total
205
179
54.15
54.82
—
—
24.42
24.85
平均 Mean
8.54
7.46
2.26
2.28
0.26
0.36
1.02
1.04
表1 24对SSR引物在全部参试资源以及微核心种质中扩增的等位变异数、有效变异数和Shannon’s信息指数 Table 1 Alleles, effective alleles, and polymorphic information of SSR amplified products in all tested genotypes and mini-corecollections of faba bean
表2 24对SSR引物在国内春播、秋播资源以及国外资源扩增的等位变异数、有效等位变异数与Shannon’s信息指数 Table 2 Alleles, effective alleles, and polymorphic information of SSR amplified products among spring, winter sowing accessions from China and accessions from abroad
SSR位点 SSR primer pair
国内春播蚕豆资源 Spring sowing accessions from China
国内秋播蚕豆资源 Winter sowing accessions from China
国外蚕豆资源 Accessions from abroad
Na
Ne
I
Na
Ne
I
Na
Ne
I
SSR6221
8
1.93
1.08
8
1.80
0.91
7
2.06
1.10
SSR5594
4
1.92
0.81
3
2.20
0.87
5
2.19
0.95
SSR2577
6
2.87
1.21
5
2.15
0.85
6
2.59
1.11
SSR6613
2
1.86
0.65
2
1.87
0.66
2
1.74
0.62
SSR1929
9
3.14
1.44
9
3.49
1.51
9
1.81
1.05
SSR3637
4
1.10
0.22
5
1.25
0.39
6
1.37
0.64
SSR5078
5
1.91
0.86
5
1.94
0.86
6
2.92
1.28
SSR2992
4
2.37
1.01
4
2.27
0.93
5
1.51
0.66
SSR9810
5
2.74
1.26
7
2.66
1.20
8
3.87
1.54
SSR10581
16
5.85
2.04
14
3.21
1.39
24
7.85
2.38
SSR4056
7
2.63
1.22
7
2.33
1.06
9
3.26
1.54
SSR10177
5
1.46
0.63
6
1.17
0.37
9
3.37
1.44
SSR3372
8
3.09
1.34
10
3.36
1.46
15
6.28
2.07
SSR1788
7
1.42
0.68
7
1.24
0.48
9
4.26
1.65
SSR5740
6
1.86
0.92
8
1.53
0.69
7
2.27
1.05
SSR4798
16
3.54
1.65
14
2.23
1.12
14
2.18
1.34
SSR3581
6
2.10
1.02
7
1.25
0.43
9
2.41
1.29
SSR2364
5
1.19
0.39
8
1.29
0.55
9
1.72
0.91
SSR6092
4
1.28
0.46
4
1.27
0.39
4
1.09
0.22
SSR4076
4
1.83
0.70
3
1.87
0.66
3
1.87
0.67
SSR6116
3
1.65
0.65
4
1.57
0.59
4
1.45
0.53
SSR102
9
2.61
1.32
8
2.03
0.99
9
3.94
1.55
SSR10296
5
1.23
0.44
6
1.82
0.84
6
2.15
1.05
SSR10421
7
1.23
0.46
8
1.36
0.62
11
1.50
0.80
合计 Total
155
52.80
—
162
47.15
—
196
65.65
—
平均 Mean
6.46
2.20
0.94
6.75
1.96
0.83
8.17
2.74
1.14
表2 24对SSR引物在国内春播、秋播资源以及国外资源扩增的等位变异数、有效等位变异数与Shannon’s信息指数 Table 2 Alleles, effective alleles, and polymorphic information of SSR amplified products among spring, winter sowing accessions from China and accessions from abroad
郑卓杰, 王述民, 宗绪晓. 中国食用豆类学. 北京: 中国农业出版社, 1997. pp93-140ZhengZ J, WangS M, ZongX X. Food Legume Crops in China. Beijing: China Agriculture Press, 1997. pp93-140(in Chinese)[本文引用:1]
[2]
FAO. Statistical Database, Food and Agriculture Organization (FAO) of the United Nations, Rome, 2012. http://faostat.fao.org/[本文引用:1]
[3]
Vand eV W, WaughR, DuncanN. Development of 8 genetic linkage map Vicia faba using molecular and biochemical techniques. Aspects Appl Biol, 199l, 27: 49-54[本文引用:1]
[4]
KlosK L E, PazM M, MarekL F, CreganP B, ShoemakerR C. Molecular markers useful for detecting resistance to brown stem rot in soybean. Crop Sci, 2000, 40: 1445-1452[本文引用:1][JCR: 1.513]
[5]
MaY, BaoS Y, YangT, HuJ G, GuanJ P, HeY H, WangX J, WanY L, SunX L, JiangJ Y, GongC X, ZongX X. Genetic linkage map of Chinese native variety faba bean (Vicia faba L. ) based on simple sequence repeat markers. Plant Breed, 2013, 132: 397-400[本文引用:2][JCR: 1.175]
[6]
ZongX X, LiuX J, GuanJ P, WangS M, LiuQ C, PaulJ G, ReddenR. Molecular variation among Chinese and global winter faba bean germplasm. Theor Appl Genet, 2009, 118: 971-978[本文引用:2][JCR: 3.658]
[7]
WangH F, ZongX X, GuanJ P, YangT, SunX L, MaY, ReddenR. Genetic diversity and relationship of global faba bean (Vicia faba L. ) germplasm revealed by ISSR markers. Theor Appl Genet, 2012, 124: 789-797[本文引用:2][JCR: 3.658]
[8]
FrankelO H, BrownA H D. Current plant genetic resources-a-critical appraisal. In: Genetics: New Frontiers Vol. IV. Oxford and IBH Publ. Co. , New Delhi, India, 1984. pp1-11[本文引用:1]
[9]
李自超, 张洪亮, 曹永生, 裘宗恩, 魏兴华, 汤圣祥, 余萍, 王象坤. 中国地方稻种资源初级核心种质取样策略研究. 作物学报, 2003, 29: 20-24LiZ C, ZhangH L, CaoY S, QiuZ E, WeiX H, TangS X, YuP, WangX K. Studies on the sampling strategy for primary core collection of Chinese ingenious rice. Acta Agron Sin, 2003, 29: 20-24 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[10]
WangL X, GuanY, GuanR X, LiY H, MaY S, DongZ M, LiuX, ZhangH Y, ZhangY Q, LiuZ X, ChangR Z, XuH M, LiL H, LinF Y, LuanW J, YanZ, NingX H, ZhuL, CuiY H, PiaoR H, LiuY, ChenP Y, QiuL J. Establishment of Chinese soybean (Glycine max) core collections with agronomic traits and SSR markers. Euphytica, 2006, 151: 215-223[本文引用:3][JCR: 1.643]
[11]
王丽侠, 李英慧, 李伟, 朱莉, 关媛, 宁学成, 关荣霞, 刘章雄, 常汝镇, 邱丽娟. 长江春大豆核心种质构建及分析. 生物多样性, 2004, 12: 578-585WangL X, LiY H, LiW, ZhuL, GuanY, NingX C, GuanR X, LiuZ X, ChangR Z, QiuL J. Establishment of a core collection of Changjiang spring sowing soybean. Biodivers Sci, 2004, 12: 578-585 (in Chinese with English abstract)[本文引用:2]
[12]
刘勇, 孙中海, 刘德春, 吴波, 周群. 利用分子标记技术选择柚类核心种质资源. 果树学报, 2006, 23: 339-345LiuY, SunZ H, LiuD C, WuB, ZhouQ. Screening the core collection of pomelo germplasm based on molecular marker. J Fruit Sci, 2006, 23: 339-345 (in Chinese with English abstract)[本文引用:1][CJCR: 1.045]
[13]
杨美, 付杰, 向巧彦, 刘艳玲. 利用AFLP分子标记技术构建花莲核心种质资源. 中国农业科学, 2011, 44: 3193-3205YangM, FuJ, XiangQ Y, LiuY L. The core-collection construction of flower lotus based on AFLP molecular markers. Sci Agric Sin, 2011, 44: 3193-3205 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[14]
EllisP R, Pink D A C, PhelpsK, JukesP L, BreedsS E, PinnegarA E. Evaluation of a core collection of Brassica oleracea accessions for resistance to Brevicoryne brassicae, the cabbage aphid. Euphytica, 1998, 103: 149-160[本文引用:1][JCR: 1.643]
[15]
MiklasP N, DelormeR, HannanR, DicksonM H. Using a sub sample of the core collection to identify new sources of resistance to white mold in common bean. Crop Sci, 1999, 39: 569-573[本文引用:1][JCR: 1.513]
[16]
SantosM R, DiasJ S. Evaluation of a core collection of Brassica oleracea accessions for resistance to white rust of crucifers (Albugo cand ida) at the cotyledon stage. GenetRes Crop Evol, 2004, 51: 713-722[本文引用:1]
[17]
DellaportaS L, WoodJ, HicksJ B. A plant DNA mini preparation: Version II. Plant Mol Biol, 1983, 1: 19-21[本文引用:1][JCR: 3.518]
[18]
DoyleJ J, DoyleJ L. Isolation of plant DNA from fresh tissue. Focus, 1990, 12: 149-151[本文引用:1]
[19]
RohlfF. NtSYS pc: Numerical Taxonomy System (Ver. 2. 2). Setauket, New York: Exeter Publishing, Ltd. ,2006[本文引用:1]
[20]
FalushD, StephensM, PritchardJ K. Inference of population structure: extensions to linked loci and correlated allele frequencies. Genetics, 2003, 164: 1567-1587[本文引用:1][JCR: 4.389]
[21]
YehF C, Boyle T J B. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg J Bot, 1997, 129: 157[本文引用:1][JCR: 0.625]
[22]
崔艳华, 邱丽娟, 常汝镇, 吕文河. 植物核心种质研究进展. 植物遗传资源学报, 2003, 4: 279-284CuiY H, QiuL J, ChangR Z, LüW H. Advances in research on core collection of plant germplasm resources. J Plant Genet Resour, 2003, 4: 279-284 (in Chinese with English abstract)[本文引用:1]
[23]
杨菁, 迟德钊, 刘玉皎, 马晓岗. 基于形态性状的青海蚕豆核心种质的初步构建. 分子植物育种, 2009, 7: 599-606YangJ, ChiD Z, LiuY J, MaX G. Preliminary construction of core collection of broad bean in Qinghai based on morphological traits. Mol Plant Breed, 2009, 7: 599-606 (in Chinese with English abstract)[本文引用:1][CJCR: 1.23]
[24]
ZongX X, ReddenR J, LiuC Q, WangS M, GuanJ P, LiuJ, XuY H, LiuX J, GuJ, YanL, AdesP, FordR. Analysis of a diverse global Pisum sp. collection and comparison to a Chinese local P. sativum collection with microsatellite markers. Theor Appl Genet, 2009, 118: 193-204[本文引用:1][JCR: 3.658]
[25]
刘玉皎, 侯万伟. 青海蚕豆种质资源AFLP多样性分析和核心资源构建. 甘肃农业大学学报, 2011, 46(4): 62-68LiuY J, HouW W. Diversity analysis on germplasm resources by AFLP and core resources construction of Vicia faba in Qinghai. J Gansu Agric Univ, 2011, 46(4): 62-68 (in Chinese with English abstract)[本文引用:1][CJCR: 0.833]
[26]
Hintum V Th J L, BothmerR V, VisserD L. Sampling strategies for composing a core collection of cultivated barley (Hordeum vulgare s. lat. ) collection in China. Hereditas, 1995, 122: 7-17[本文引用:1][CJCR: 0.928]
[27]
BalfourierF, CharmetG, ProsperiJ M, GoulardM, MonestiezP. Comparison of different spatial strategies for sampling a core collection of natural populations of folder crops. Genet SelEvol, 1998, 30: 215-235[本文引用:1]
[28]
OrtizR, Ruiz-TapiaF N, Mujica-SanchezA. Sampling strategy for a core collection of Peruvian quinoa germplasm. Theor Appl Genet, 1998, 96: 475-483[本文引用:1][JCR: 3.658]
[29]
GizlicelZ. Genetic base for North American public soybean cultivars released between 1947-1988. Crop Sci, 1994, 34: 1143-1147[本文引用:1][JCR: 1.513]
[30]
MalosettiM, AbadieT. Sampling strategy to develop a core collection of Uruguayan maize land races based on morphological traits. Genet ResCrop Evol, 2001, 48: 381-390[本文引用:1][JCR: 1.593]
[31]
Chand raS, HuamanZ, KrishnaH S, OrtizR. Optimal sampling strategy and core collection size of Andean tetraploid potato based on isozyme data: a simulation study. Theor Appl Genet, 2002, 104: 1325-1334[本文引用:1][JCR: 3.658]

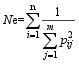



{kind=link}
{kind=link}