关键词:甘蓝型油菜; 单核苷酸多态性; 发芽率; 盐胁迫; 干旱胁迫; 数量性状座位 QTL Mapping for Germination Percentage under Salinity and Drought Stresses inBrassica napus L. Using a SNP Genetic Map JIAN Hong-Ju**, XIAO Yang**, LI Jia-Na, MA Zhen-Zhen, WEI Li-Juan, LIU Lie-Zhao* College of Agronomy and Biotechnology, Southwest University / Chongqing Engineering Research Center for Rapeseed, Chongqing 400716, China Fund: AbstractThe objective of this study was to identify QTLs for seed germination percentage under salinity and drought stresses inBrassica napus L. using a recombinant inbred line (RIL) population derived from the cross between yellow-seeded GH06 (female parent) and black-seeded P174 (male parent). The RILs were F11 developed via single-seed-descent propagation. QTLs for germination percentage under 16 g L-1 NaCl and 20% PEG-6000 (w/w) treatment, after four-day treatment and on the third and fourth day, were detected using the composite interval mapping (CIM) method. The linkage map contains 2795 SNP markers with the total length of 1832.9 cM and the average distance of 0.66 cM. A total of 19 QTLs were mapped on chromosomes A01, A03, A06, A07, A09, and C06. Twelve of them were identified under salinity stress with explained phenotypic variation from 4.9% to 10.9%, and eight QTLs were related to drought stress with explained phenotypic variation from 3.8% to 6.9%. The QTLs on A03 and A09 were located in close marker regions. Our results showed that the germination percentage of rapeseed is controlled by many minor-effect loci with great influence by environmental factors. Different genes are triggered in response to salinity or drought stress along with the stress duration.
Keyword:Brassica napus; SNP; Germination percentage; Salinity stress; Drought stress; QTL Show Figures Show Figures
表1 亲本和重组自交系群体在不同胁迫环境下的发芽率 Table 1 Germination percentage under different stress environments in parents and RIL population
胁迫环境 Stress environment
亲本Parent
重组自交系群体 RIL population
GH06 (%)
P174 (%)
均值 Mean (%)
标准差 SD (%)
范围 Range (%)
方差 Variance
偏度 Skewness
峰度 Kurtosis
变异系数 CV
S4
83.33
48.33
66.74
32.18
0-100.00
1035.51
-0.68
-0.91
0.48
SS3
25.00
29.17
33.95
24.44
0-91.67
597.25
0.39
-0.91
0.72
SS4
51.67
16.67
23.88
19.16
0-88.33
367.24
0.79
0.09
0.80
D4
99.17
92.50
79.66
24.47
0-100.00
598.91
-1.51
1.52
0.31
DD3
13.33
53.33
45.95
22.82
0-100.00
520.96
-0.08
-0.67
0.50
DD4
78.33
37.50
19.40
15.96
0-76.67
254.77
0.88
0.25
0.82
S4: 盐胁迫累计4 d; SS3: 盐胁迫第3天; SS4: 盐胁迫第4天; D4: 干旱胁迫累计4 d; DD3: 干旱胁迫第3天; DD4: 干旱胁迫第4天。 S4: 4 days under salinity stress; SS3: the third day under salinity stress; SS4: the fourth day under salinity stress; D4: 4 days under drought stress; DD3: the third day under drought stress; DD4: the fourth day under drought stress.
表1 亲本和重组自交系群体在不同胁迫环境下的发芽率 Table 1 Germination percentage under different stress environments in parents and RIL population
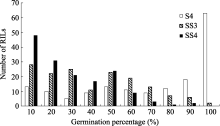
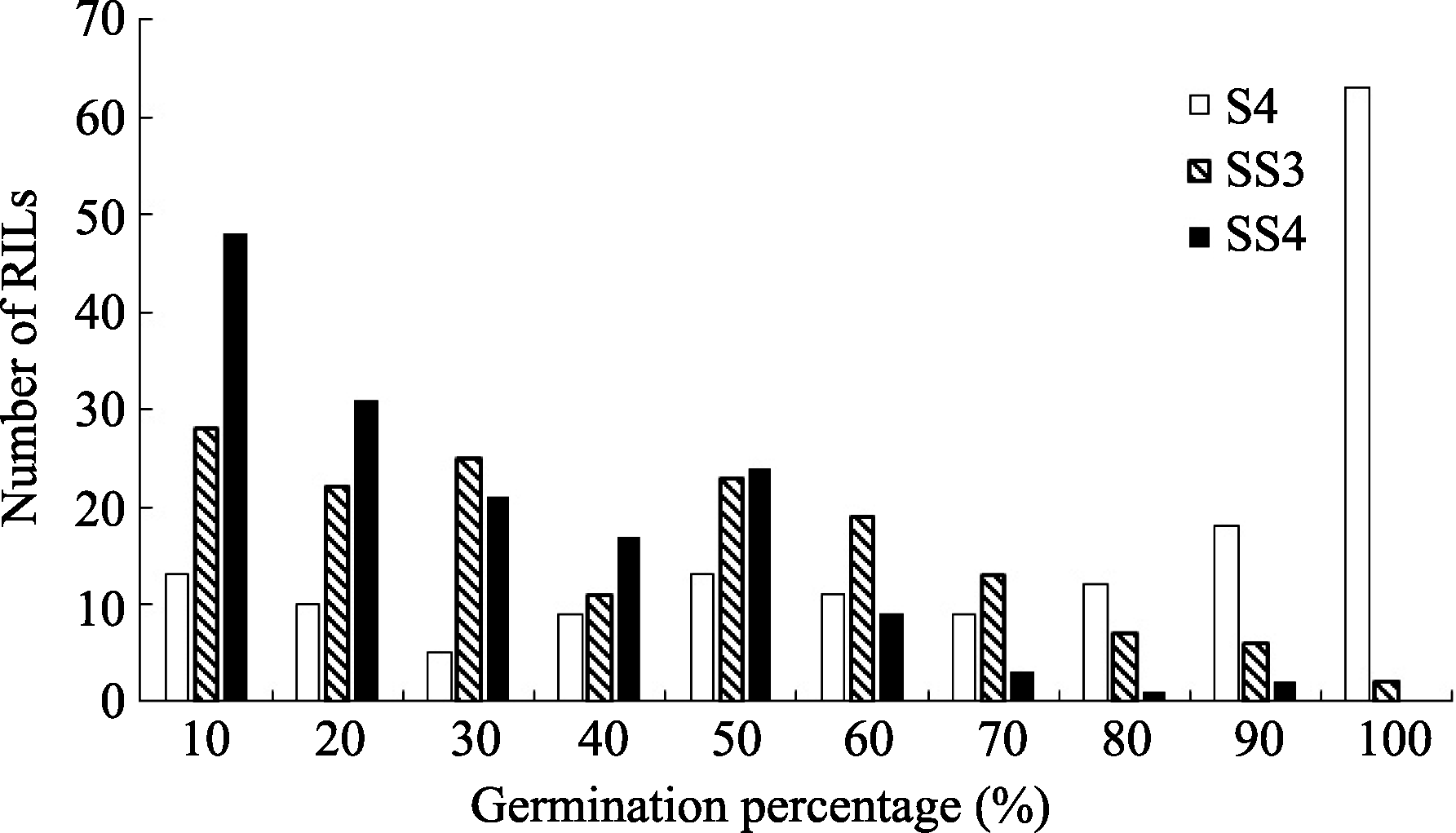
图1 盐胁迫下重组自交系群体发芽率的频率分布S4: 盐胁迫累计4 d; SS3和SS4: 盐胁迫第3和第4天。Fig. 1 Frequency distribution of the germination percentage of RILs under salinity stressS4: 4 days under salinity stress; SS3 and SS4: the third and fourth day under salinity stress.
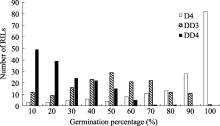
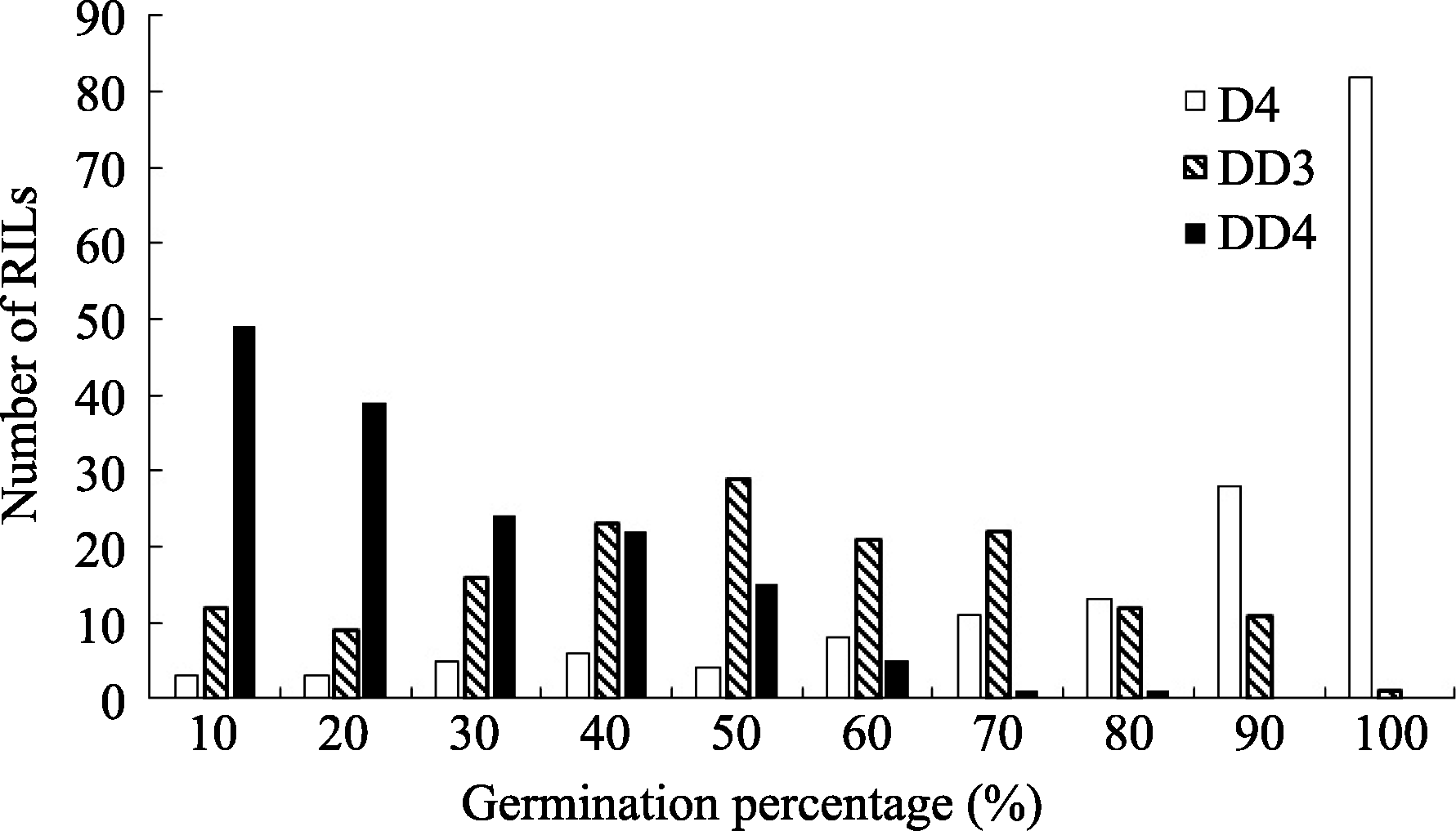
图2 干旱胁迫下重组自交系群体发芽率的频率分布D4: 干旱胁迫累计4 d; DD3和DD4: 干旱胁迫第3和第4天。Fig. 2 Frequency distribution of the germination percentage of the RILs under drought stressD4: 4 days under drought stress; DD3 and DDR: the third and fourth day under drought stress.
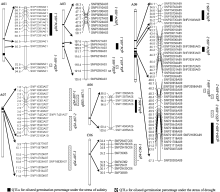
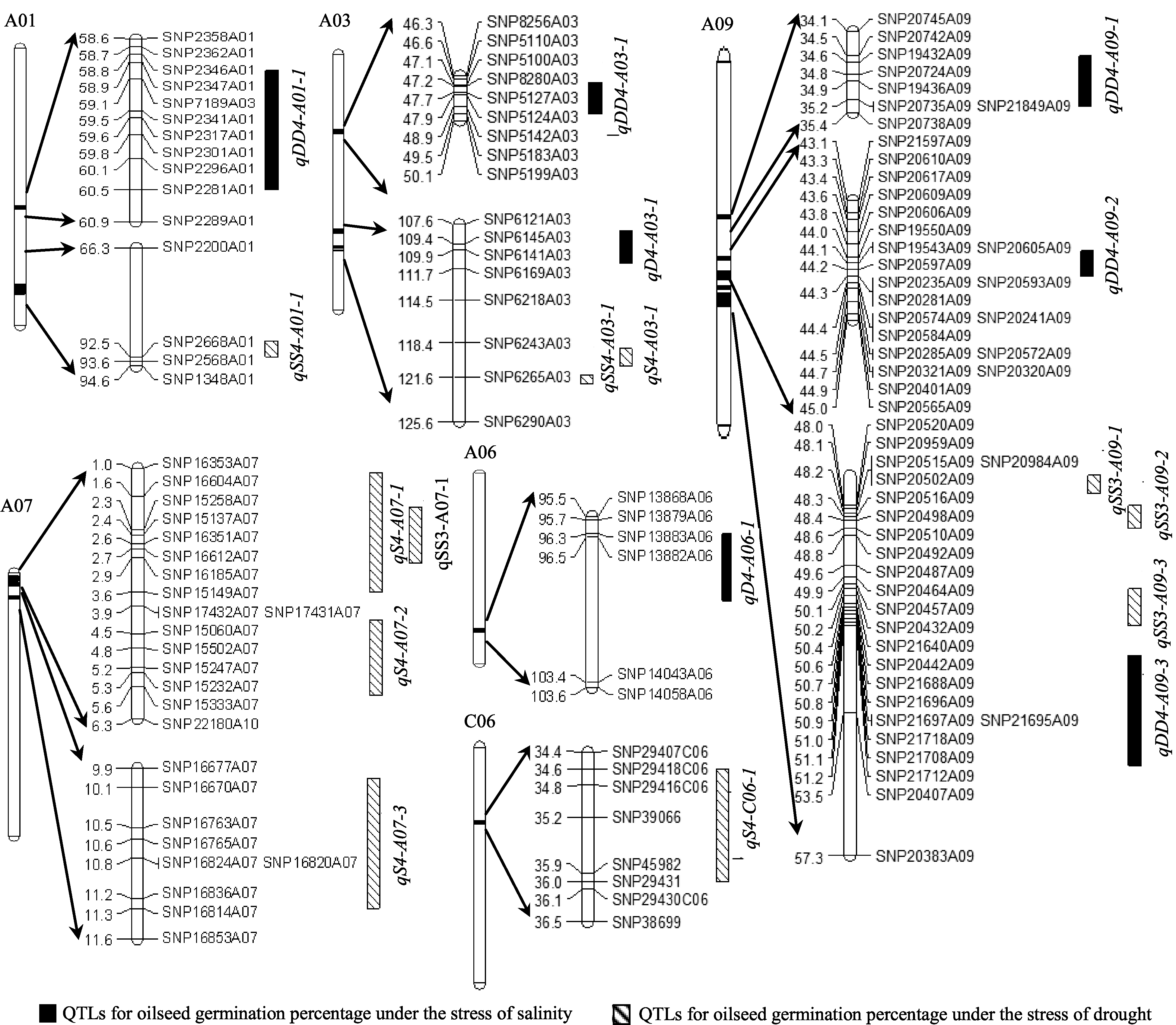
图3 甘蓝型油菜种子发芽率与盐胁迫、干旱胁迫相关的QTL在遗传图谱的分布Fig. 3 Putative QTLs of oilseed germination percentage under salinity and drought stresses in the genetic linkage map
傅廷栋. 中国油菜生产和品种改良的现状与前景. 安徽农学通报, 2000, 6(1): 2-8FuT D. Chinese rapeseed production and the improvement of the status and prospects. Anhui Agric Sci Bull, 2000, 6(1): 2-8 (in Chinese with English abstract)[本文引用:1][CJCR: 0.2433]
[2]
刘祖祺, 张石城. 植物抗性生理学. 北京: 中国农业出版社, 1994. pp222-223LiuZ Q, ZhangS C. Plant Resistance Physiology. Beijing: China Agriculture Press, 1994. pp222-223(in Chinese)[本文引用:1]
[3]
戴清明, 吕爱钦, 何维君, 谢年保, 陈欣, 张志远, 匡朝凌, 瞿科. 洞庭湖区油菜主要气象灾害发生规律与减灾避灾对策. 作物研究, 2006, (1): 60-63DaiQ M, LüA Q, HeW J, XieN B, ChenX, ZhangZ Y, KuangC L, QuK. Thearising regulation and the decreasing and avoiding strategies of the main meteorological disasters on rapeseed in Dongting lake region. Crop Res, 2006, (1): 60-63 (in Chinese with English abstract)[本文引用:1][CJCR: 0.7044]
[4]
LombardV, DelourmeR. A consensus linkage map for rapeseed (Brassica napus L. ): construction and integration of three individual maps from DH populations. Theor Appl Genet, 2001, 103: 491-507[本文引用:1][JCR: 3.658]
[5]
EckeW, UzunovaM, WeisslederK. Mapping the genome of rapeseed (Brassica napus L. ): II. Localization of genes controlling erucic acid synthesis and seed oil content. Theor Appl Genet, 1995, 91: 972-977[本文引用:1][JCR: 3.658]
[6]
ThormannC E, RomeroJ, MantetJ, OsbornT C. Mapping loci controlling the concentrations of erucic and linolenic acids in seed oil of Brassica napus L. Theor Appl Genet, 1996, 93: 282-286[本文引用:2][JCR: 3.658]
[7]
ToroserD, ThormannC E, OsbornT C, MithenR. RFLP mapping of quantitative trait loci controlling seed aliphatic- glucosinolate content in oilseed rape (Brassica napus L). Theor Appl Genet, 1995, 91: 802-808[本文引用:2][JCR: 3.658]
[8]
HowellP M, SharpeA G, LydiateD J. Homoeologous loci control the accumulation of seed glucosinolates in oilseed rape (Brassica napus). Genome Res, 2003, 46: 454-460[本文引用:2][JCR: 14.397]
[9]
ZhaoJ Y, BeckerH C, ZhangD Q, ZhangY F, EckeW. Conditional QTL mapping of oil content in rapeseed with respect to protein content and traits related to plant development and grain yield. Theor Appl Genet, 2006, 113: 33-38[本文引用:1][JCR: 3.658]
[10]
ZhaoJ W, MengJ L. Genetic analysis of loci associated with partial resistance to Sclerotinia sclerotiorum in rapeseed (Brassica napus L. ). Theor Appl Genet, 2003, 106: 759-764[本文引用:1][JCR: 3.658]
[11]
FooladM R, LinG Y, ChenF Q. Comparison of QTLs for seed germination under non-stress, cold stress and salt stress in tomato. Plant Breed, 1999, 118: 167-173[本文引用:1][JCR: 1.175]
[12]
BetteyM, Finch-SavageW E, KingG J, LynnJ R. Quantitative genetic analysis of seed vigour and pre-emergence seedling growth traits in Brassica oleracea. New Phytol, 2000, 148: 277-286[本文引用:1][JCR: 6.736]
[13]
KrishnasamyV, SeshuD V. Seed germination rate and associated characters in rice. Crop Sci, 1989, 29: 904-908[本文引用:1][JCR: 1.513]
[14]
许耀照, 孙万仓, 曾秀存, 李彩霞, 周喜旺. 盐碱胁迫冬油菜的主导因素分析. 草业科学, 2013, (3): 423-429XuY Z, SunW C, ZengX C, LiC X, ZhouX W. Analysis of the key stress factors in winter rape under simulated salinity- alkalinity mixed condition. Pratac Sci, 2013, (3): 423-429 (in Chinese with English abstract)[本文引用:1]
[15]
原小燕, 符明联, 何晓莹. 不同抗旱性油菜种子萌发期抗旱指标比较研究. 干旱地区农业研究, 2012, 30(5): 77-81YuanX Y, FuM L, HeX Y. The comparative study on drought resistance index of rape with different drought resistance in germination. Agric Res Arid Areas, 2012, 30(5): 77-81 (in Chinese with English abstract)[本文引用:1]
[16]
谢小玉, 张兵, 陈思岑. 油菜发芽期和苗期抗旱性鉴定评价方法. 农机化研究, 2013, (2): 112-116XieX Y, ZhangB, ChenS C. Method of identification for characteristics of drought-against on germination and seedling growth of rape materials. J Agric Mechaniz Res, 2013, (2): 112-116 (in Chinese with English abstract)[本文引用:1]
[17]
NguyenT, FriedtW, SnowdonR. Cloning and mapping of a cand idate gene for germination and seedling vigour in yellow-seeded oilseed rape. In: Fu T D ed. The 12th International Rapeseed Congress, Wuhan, China, 2007. p4[本文引用:1]
[18]
刘丹, 刘贵华, 王汉中. 油菜抗性相关基因的分离及其基因工程研究进展. 中国农业科技导报, 2008, 8(3): 6-11LiuD, LiuG H, WangH Z. Isolation of resistant genes and their gene engineering research development in rapeseed. J Agric Sci Technol, 200, 8(3): 6-11 (in Chinese with English abstract)[本文引用:1]
[19]
LiuL, QuC, WittkopB, YiB, XiaoY, HeY, SnowdonR J, LiJ. A high-density SNP map for accurate mapping of seed fibre QTL in Brassica napus L. PLoS One, 2013, 8(12): e83052. DOI: 10.1371/journal.pone.0083052[本文引用:1][JCR: 3.73]
[20]
WangS, BastenC J, ZengZ B. Windows QTL Cartographer. Ver. 2. 5 [computer program] Department of Statistics, North Carolina State University, Raleigh, NC, 2006. http://statgen.ncsu.edu/qtlcart/WQTLCart.htm[本文引用:1]
[21]
Land erE S, BotsteinD. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics, 1989, 121: 185-199[本文引用:1][JCR: 4.389]
[22]
MccouchS R, ChoY G, YanoM, PaulE, BlinstrubM, MorishimaH, KinoshitaT. Report on QTL nomenclature. Rice Genet Newsl, 1997, 14[本文引用:1]
[23]
GanalM W, AltmannT, RoderM S. SNP identification in crop plants. Curr Opin Plant Biol, 2009, 12: 211-217[本文引用:1][JCR: 8.455]
[24]
McnallyK L, ChildsK L, BohnertR, DavidsonR M, ZhaoK, UlatV J, ZellerG, ClarkR M, HoenD R, BureauT E, StokowskiR, BallingerD G, FrazerK A, CoxD R, PadhukasahasramB, BustamanteC D, WeigelD, MackillD J, BruskiewichR M, RatschG, BuellC R, LeungH, LeachJ E. Genomewide SNP variation reveals relationships among land races and modern varieties of rice. Proc Natl Acad Sci USA, 2009, 106: 12273-12278[本文引用:1][JCR: 9.737]
[25]
RafalskiA. Applications of single nucleotide polymorphisms in crop genetics. Curr Opin Plant Biol, 2002, 5: 94-100[本文引用:1][JCR: 8.455]
[26]
孙玉燕, 刘磊, 郑峥, 张春芝, 周龙溪, 宗园园, 李涛, 李君明. 番茄耐旱和耐盐遗传改良的研究进展及展望. 园艺学报, 2012, 39: 2061-2074SunY Y, LiuL, ZhengZ, ZhangC Z, ZhouL X, ZongY Y, LiT, LiJ M. A review and perspectives on genetic improvement of salt and drought tolerance in tomato. Acta Hortic Sin, 2012, 39: 2061-2074 (in Chinese with English abstract)[本文引用:1][CJCR: 1.194]
[27]
郝岗平, 吴忠义, 陈茂盛, 曹鸣庆, BrunnelD, PelletierG, 黄丛林, 杨清. 拟南芥CBF4基因位点的单核苷酸多态性(SNP)变化与抗旱表型的相应性. 农业生物技术学报, 2004, 12: 122-131HaoG P, WuZ Y, ChenM S, CaoM Q, BrunnelD, PelletierG, HuangC L, YangQ. Single nucleotide polymorphisms of CBF4 locus region of Arabidopsis thaliana correspond to drought tolerance. J Agric Biotechnol, 2004, 12: 122-131 (in Chinese with English abstract)[本文引用:1]
[28]
VersluesP E, AgarwalM, Katiyar-AgarwalS, ZhuJ H, ZhuJ K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J, 2006, 45: 523-539[本文引用:1][JCR: 6.582]


{kind=link}
{kind=link}
{kind=link}