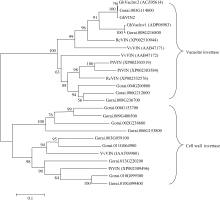
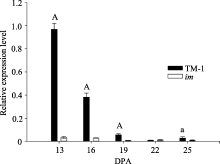
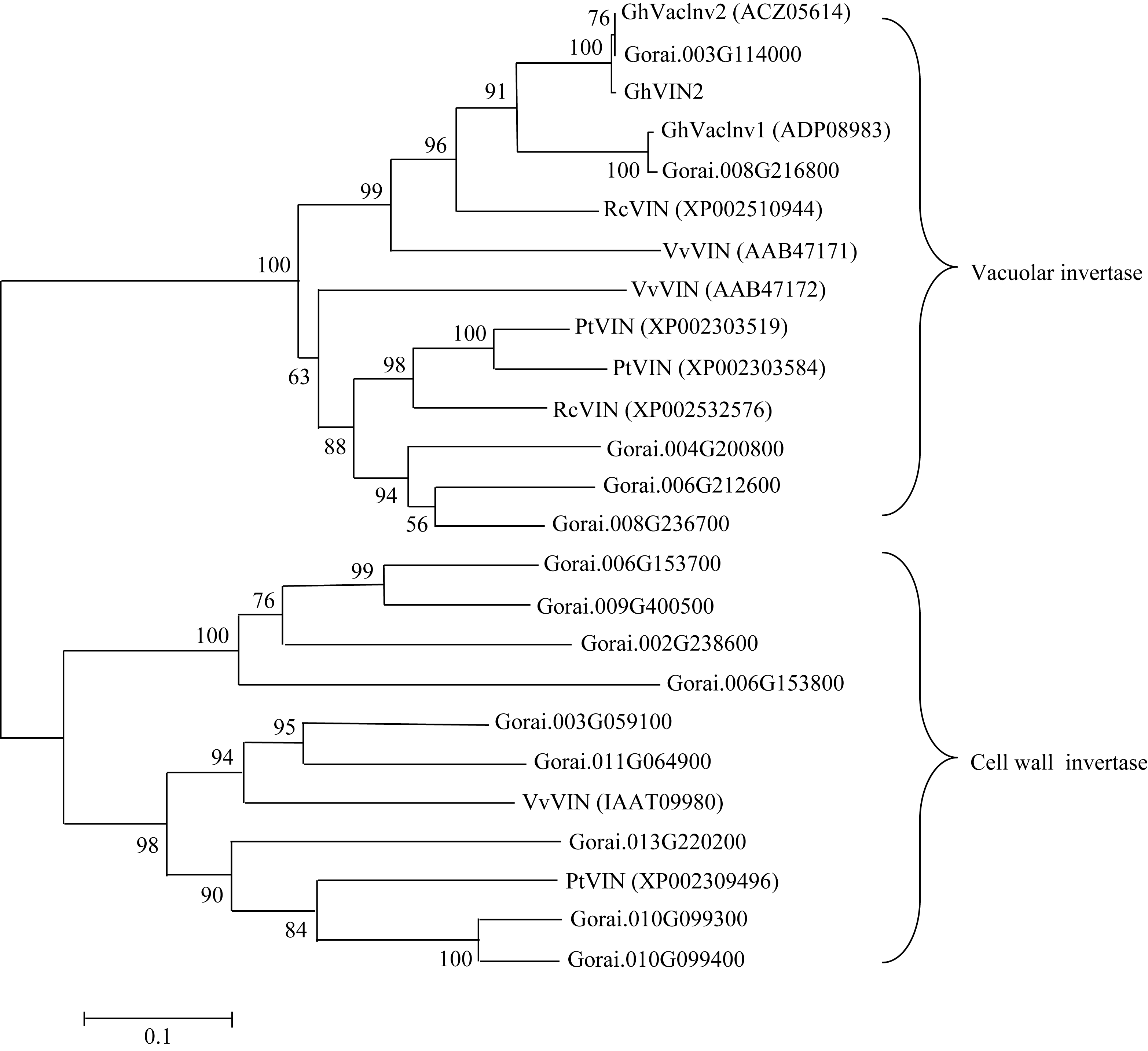
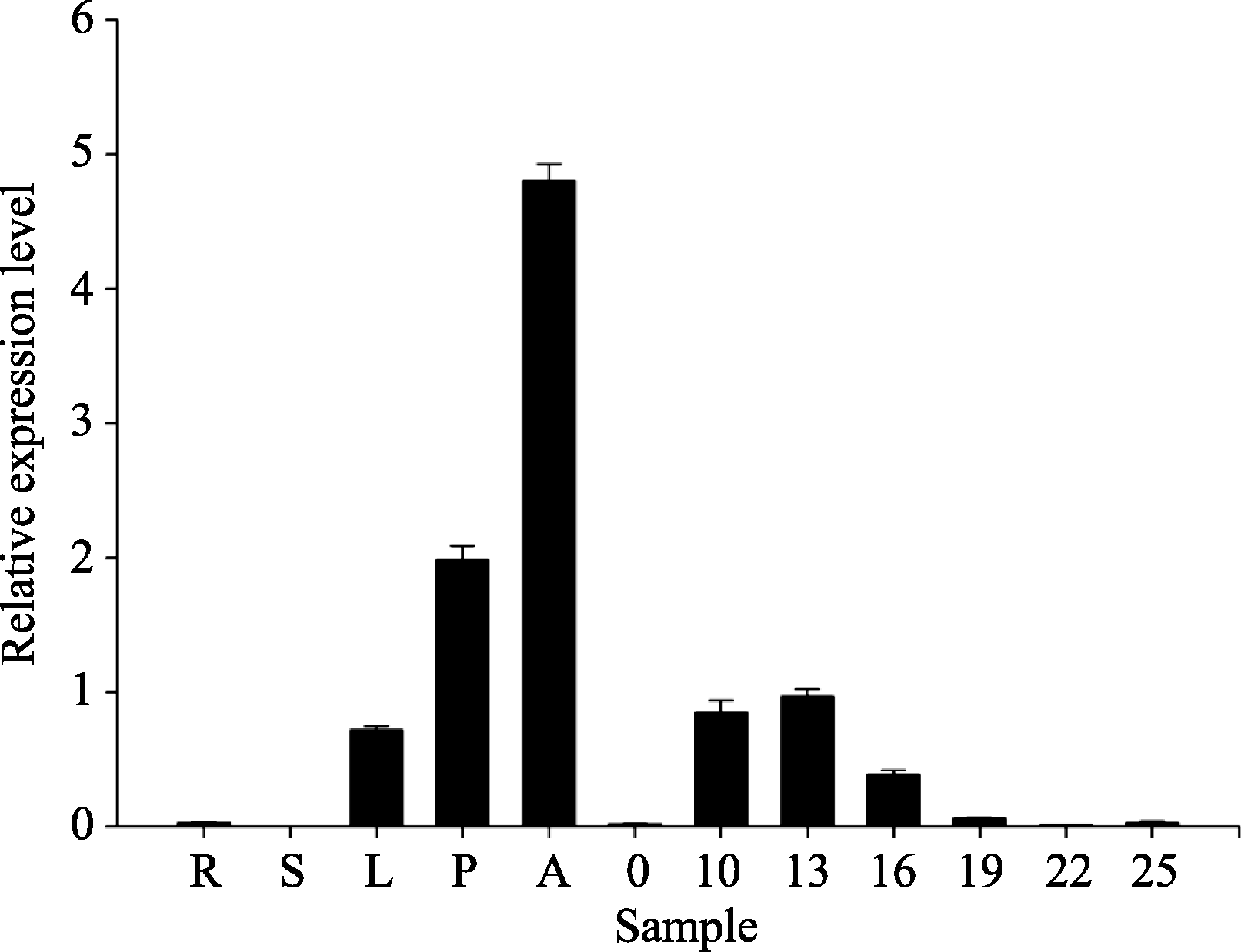
关键词:棉花; 纤维发育; 液泡转化酶2; 克隆; 功能分析 Cloning and Functional Analysis ofGhVacInc2a Encoding Vacuolar Invertase in Cotton XU Wen-Ting, WANG Cheng, XU Xiao-Yang, NIU Er-Li, CAI Cai-Ping, GUO Wang-Zhen* State Key Laboratory of Crop Genetics and Germplasm Enhancement / Hybrid Cotton R&D Engineering Research Center, Ministry of Education, Nanjing Agricultural University, Nanjing 210095, China Fund: AbstractCotton fiber development depends on a large supply of sugar. Invertase plays a central role in sucrose metabolism. Structural and functional analysis of gene encoding invertase will conductively reveal the complex molecular mechanism of fiber development, and also provide elite gene resources for improvement of cotton fiber quality. In the study, a novel gene encoding vacuolar invertase was obtained by sillico cloning, based on a differentially expressed EST (GenBank accession number EY196825) betweenG. hirsutum acc. TM-1 andim mutant, combined with reconfirmation of ORF information in transcriptional and genomic level. BlastN showed that the gene shared 99% identity ofGhVacInv2reported previously (GenBank accession number FJ864677), and designated asGhVacInv2a (GenBank accession number KF305322).GhVacInv2a contained an open reading frame of 1857 bp that encoded a polypeptide of 618 amino acids, with seven exons and six introns at genome sequence.GhVacInv2a comprised one copy in diploid cotton speciesG. herbaceum andG. raimondii, and two copies in tetraploid cotton speciesG. hirsutum acc. TM-1 andG. barbadense cv. Hai 7124, withGhVacInv2ain A-subgenome andGhVacInv2 in D-subgenome, respectively. Q-PCR expression analysis showed that the set of homoelogous genes performed the highest expressive abundance in anther in comparison with other tissues. In developing fibers, it was predominantly expressed from fiber rapid elongation stage, and there existed significant difference between TM-1 andim mutant during 13 to 19 days post anthesis (DPA) in fiber tissues. Further,GhVacInv2a was located on chromosome 3 by developing subgenome-specific SNP marker, and the association analysis showed that there was a significant correction (P=0.0087) betweenGhVacInv2a and fiber strength, suggesting thatGhVacInv2aprobably plays a key role in fiber qualities formation.
Keyword:Cotton; Fiber development; Vacuolar invertase 2; Cloning; Functional analysis Show Figures Show Figures
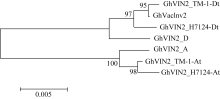
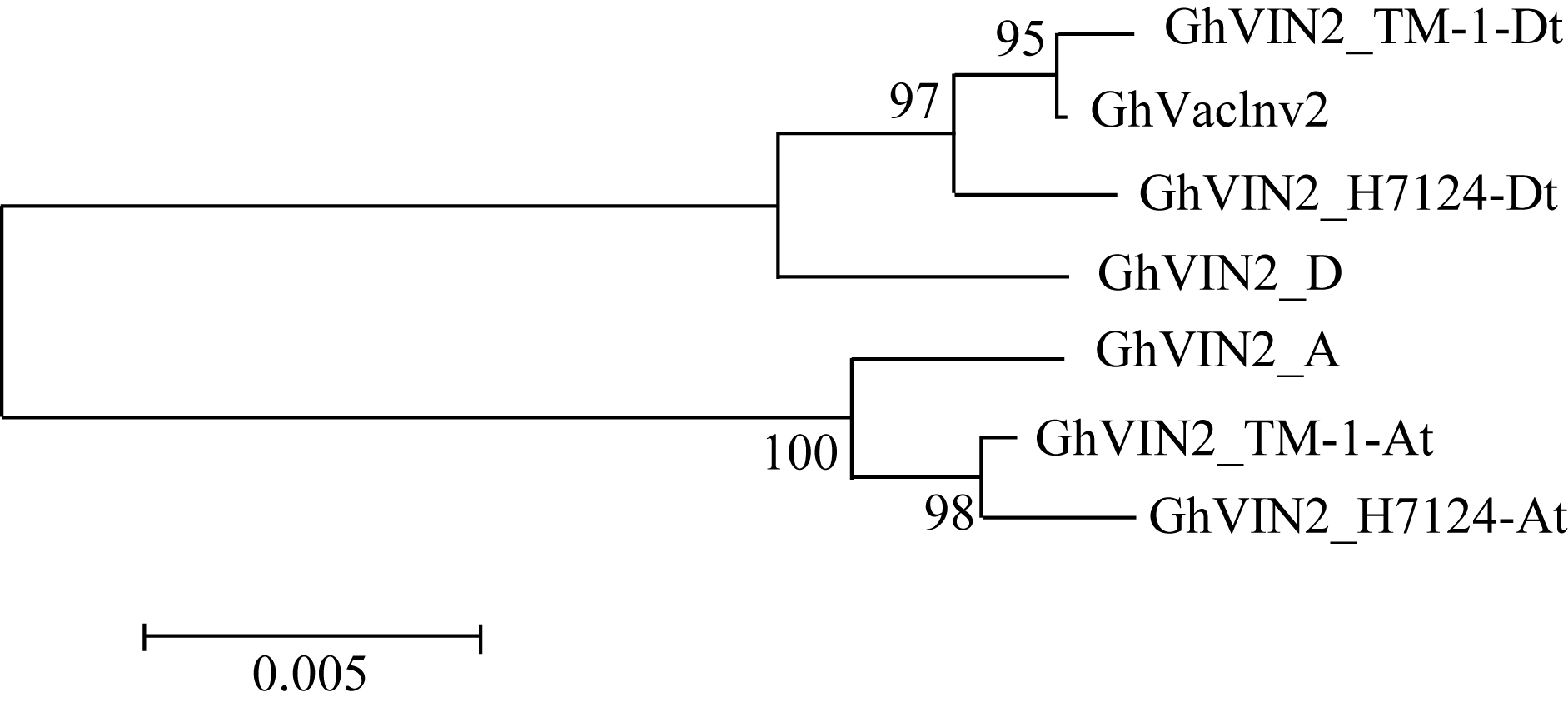
图1 GhVacInv2a类同源基因在不同棉种中的系统进化A: 阿非利加棉; D: 雷蒙德氏棉; TM-1_At: TM-1的A亚组; TM-1_Dt: TM-1的D亚组; H7124_At: 海7124的A亚组; H7124_Dt: 海7124的D亚组。GhVIN2代表 GhVacInv2a。Fig. 1 Phylogenetic analysis for GhVacInv2aorthologs in four cotton speciesA: G. herbaceum L. var. africanum; D: G. raimondii; TM-1_At: A subgenome of G. hirsutumacc.TM-1; TM-1_Dt: D subgenome of G. hirsutumacc.TM-1; H7124_At: A subgenome of G. barbadense cv. Hai7124; H7124_Dt: D subgenome of G. barbadense cv. Hai7124. GhVIN2: abbrevation for GhVacInv2a.
SturmA, TangG Q. The sucrose-cleaving enzymes of plants and crucial for development, growth and carbon partitioning. Trends Plant Sci, 1999, 4: 401-407[本文引用:2][JCR: 11.808]
[2]
RoitschT, GonzálezM C. Function and regulation of plant invertases: sweet sensations. Trends Plant Sci, 2004, 9: 606-613[本文引用:1][JCR: 11.808]
[3]
DaviesC, RobinsonS P. Sugar accumulation in grape berries: cloning of two putative vacuolar invertase cDNAs and their expression in grapevine tissues. Plant Physiol, 1996, 111: 275-283[本文引用:1][JCR: 6.555]
[4]
GreinerS, RauschT, SonnewaldU, HerbersK. Ectopic expression of a tobacco invertase inhibitor homolog prevents cold-induced sweetening of potato tubers. Nat Biotechnol, 1999, 17: 708-711[本文引用:2][JCR: 32.438]
[5]
KlannE M, ChetelatR T, BennettA B. Expression of acid invertase gene controls sugar composition in tomato (Lycopersicon) fruit. Plant Physiol, 1993, 103: 863-870[本文引用:1][JCR: 6.555]
[6]
RuanY L, JinY, YangY J, LiG J, BoyerJ S. Sugar input, metabolism, and signaling mediated by invertase: roles in development, yield potential, and response to drought and heat. Mol Plant, 2010, 3: 942-955[本文引用:2][JCR: 6.126][CJCR: 0.9487]
[7]
KohornB D, KobayashiM, JohansenS, RieseJ, HuangL F, KochK, FuS, DotsonA, ByersN. An Arabidopsis cell wall-associated kinase required for invertase activity and cell growth. Plant J, 2006, 46: 307-316[本文引用:1][JCR: 6.582]
[8]
RoitschT, GonzálezM C. Function and regulation of plant invertases: sweet sensations. Trends Plant Sci, 2004, 9: 606-613[本文引用:1][JCR: 11.808]
BasraA S, MalikC. Development of the cotton fiber. Int Rev Cytol, 1984, 89: 65-113[本文引用:1]
[11]
KimH J, TriplettB A. Cotton fiber growth in planta and in vitro. Models for plant cell elongation and cell wall biogenesis. Plant Physiol, 2001, 127: 1361-1366[本文引用:1][JCR: 6.555]
[12]
WangL, LiX R, LianH, NiD A, HeY K, ChenX Y, RuanY L. Evidence that high activity of vacuolar invertase is required for cotton fiber and Arabidopsis root elongation through osmotic dependent and independent pathways, respectively. Plant Physiol, 2010, 154: 744-756[本文引用:2][JCR: 6.555]
[13]
TaliercioE, SchefflerJ, SchefflerB. Characterization of two cotton (Gossypium hirsutum L. ) invertase genes. Mol Biol Rep, 2010, 37: 3915-3920[本文引用:2][JCR: 2.506]
武耀廷, 刘进元. 一种高效提取棉花不同组织总RNA的热硼酸改良法. 棉花学报, 2004, 16(2): 67-71WuY T, LiuJ Y. A modified hot borate method for efficient isolation of total RNA from different cotton tissues. Cotton Sci, 2004, 16(2): 67-71 (in Chinese with English abstract)[本文引用:1][CJCR: 1.462]
[16]
蒋建雄, 张天真. 利用CTAB/酸酚法提取棉花组织总RNA. 棉花学报, 2003, 15: 166-167JiangJ X, ZhangT Z. Extraction of total RNA in cotton tissues with CTAB-acidic phenolic method. Cotton Sci, 2003, 15: 166-167 (in Chinese with English abstract)[本文引用:1][CJCR: 1.462]
[17]
PatersonA H, BrubakerC L, JonathanF. Rapid method for extraction of cotton (Gossypium spp. ) genomic DNA suitable for RFLP or PCR analysis. Plant Mol Biol Rep, 1993, 11: 122-127[本文引用:1][JCR: 5.319]
[18]
PfafflM W. A new mathematical model for relative quantification in real-time RT-PCR. Nucl Acids Res, 2001, 29: e45[本文引用:1]
[19]
ZhaoL, LvY D, CaiC P, TongX C, ChenX D, ZhangW, DuH, GuoX H, GuoW Z. Toward allotetraploid cotton genome assembly: integration of a high-density molecular genetic linkage map with DNA sequence information. BMC Genomics, 2012, 13: 539[本文引用:2][JCR: 4.397]
[20]
WangC, ZhangT Z, GuoW Z. The im mutant gene negatively affects many aspects of fiber quality traits and lint percentage in cotton. Crop Sci, 2013, 53: 27-37[本文引用:3][JCR: 1.513]
[21]
RossH A, DaviesH V, BurchL R, ViolaR, McRaeD. Developmental changes in carbohydrate content and sucrose degrading enzymes in tuberising stolons of potato (Solanum tuberosum). Physiol Plant, 1994, 90: 748-756[本文引用:2][JCR: 6.555]
[22]
TangG Q, LuscherM, SturmA. Antisense repression and vacuolar and cell wall invertase in transgenic carrot alters early plant development and sucrose partitioning. Plant Cell, 1999, 11: 177-189[本文引用:2][JCR: 9.251]
[23]
AndersenM N, AschF, WuY, JensenC R, NaestedH, MogensenV O, KochK E. Soluble invertase expression is an early target of drought stress during the critical, abortion-sensitive phase of young ovary development in maize. Plant Physiol, 2002, 130: 591-604[本文引用:1][JCR: 6.555]
[24]
KlannE M, HallB, BennettA B. Antisense acid invertase (TIV1) gene alters soluble sugar composition and size in transgenic tomato fruit. Plant Physiol, 1996, 112: 1321-1330[本文引用:1][JCR: 6.555]
[25]
DaviesC, RobinsonS P. Sugar accumulation in grape berries: cloning of two putative vacuolar invertase cDNAs and their expression in grapevine tissues. Plant Physiol, 1996, 111: 275-283[本文引用:1][JCR: 6.555]
[26]
YauY, SimonP. A 2. 5-kb insert eliminates acid soluble invertase isozyme II transcript in carrot (Daucus carota L. ) roots, causing high sucrose accumulation. Plant Mol Biol, 2003, 53: 151-162[本文引用:1][JCR: 3.518]
[27]
SergeevaL I, Keurentjes J J B, BentsinkL, VonkJ, van der Plas L H W, KoornneefM, VreugdenhilD. Vacuolar invertase regulates elongation of Arabidopsis thaliana roots as revealed by QTL and mutant analysis. Proc Nat Acad Sci USA, 2006, 103: 2994-2999[本文引用:1]
[28]
LongJ C, ZhaoW, RashotteA M, MudayG K, HuberS C. Gravity-stimulated changes in auxin and invertase gene expression in maize pulvinal cells. Plant Physiol, 2002, 128: 591-602[本文引用:1][JCR: 6.555]
[29]
TrouverieJ, ThévenotC, RocherJ P, SottaB, PrioulJ L. The role of abscisic acid in the response of a specific vacuolar invertase to water stress in the adult maize leaf. J Exp Bot, 2003, 54: 2177-2186[本文引用:1][JCR: 5.242]
[30]
MitsuhashiW, SasakiS, KanazawaA, YangY Y, KamiyaY, ToyomasuT. Differential expression of acid invertase genes during seed germination in Arabidopsis thaliana. Biosci Biotechnol Biochem, 2004, 68: 602-608[本文引用:1]
[31]
TrouverieJ, Chateau-JoubertS, ThévenotC, JacquemotM P, PrioulJ L. Regulation of vacuolar invertase by abscisic acid or glucose in leaves and roots from maize plantlets. Planta, 2004, 219: 894-905[本文引用:2][JCR: 3.347]
[32]
LiZ, PalmerW M, MartinA P, WangR, RainsfordF, JinY, PatrickJW, YangY, RuanY L. High invertase activity in tomato reproductive organs correlates with enhanced sucrose import into, and heat tolerance of, young fruit. J Exp Bot, 2012, 63: 1155-1166[本文引用:1][JCR: 5.242]
[33]
XuJ, AvigneW T, McCartyD R, KochK E. A similar dichotomy of sugar modulation and developmental expression affects both paths of sucrose metabolism: evidence from a maize invertase gene family. Plant Cell, 1996, 8: 1209-1220[本文引用:1][JCR: 9.251]

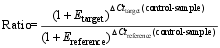


{kind=link}
{kind=link}
{kind=link}
{kind=link}