关键词:烟草; 表达序列标签; 抗病基因同源物; SSR Identification of Expressed Resistance Gene Analogues (RGAs) and Development of RGA-SSR Markers inNicotiana YUAN Qing-Hua, XIE Rui-Hong, ZHANG Zhen-Chen, MA Zhu-Wen, LI Ji-Qin, LI Shu-Ling, CHEN Jun-Biao* Crops Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China
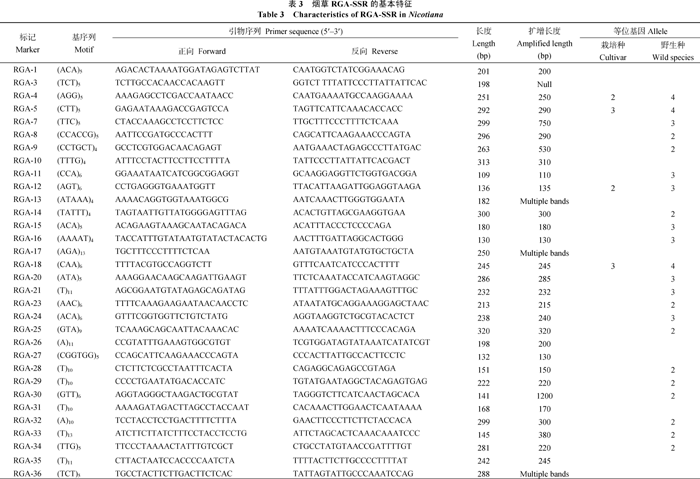
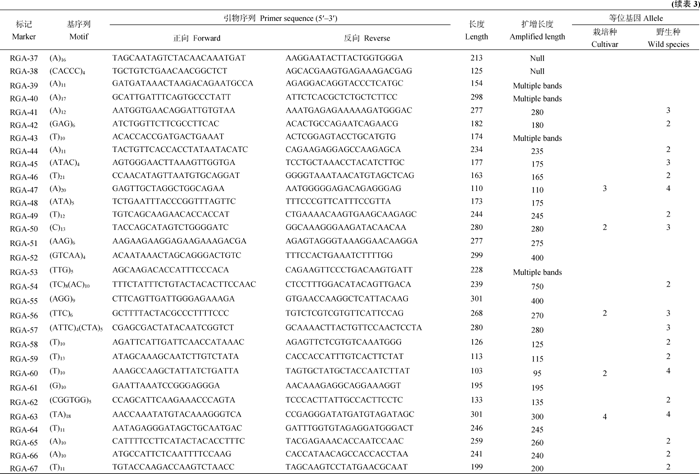
AbstractTobacco is an ideal experimental system for studing on plant-pathogen interaction. Identification of tobaccoR gene and resistance gene analogs is propitious to elucidating the underlying resistant mechanisms. In recent years, the growing public tobacco EST data provide rich source for identifying expressed RGA. In this study, 149 606 Uni-EST were assembled from 412 325 ESTs of tobacco in GenBank. By scanning the Uni-EST with 112 plant R gene protein sequences 1113 NtRGAs were identified. These expressed RGAs comprised 273, 546, 53, 102, and 30 of NBS-LRR, LRR-PK, LRR, PK, and Mlo domains encodingR genes, respectively. No domain was detected in the rest of 109 RGAs. By aligning sequence 1079 NtRGAs were allocated on 712 loci inN. benthamianagenome. A total of 78 simple sequence repeats (SSRs) were identified from 72 NtRGAs. Sixty-four primer pairs were designed base on the flanking sequence of SSR. Among them, 54 primer pairs were amplified with clear bands from tobacco genomic DNA. Nine primer pairs were detected to have polymorphism among 24 varieties ofNicotiana tabacum with two to four alleles (on average 2.56 alleles). Forty-one primer pairs were detected to have polymorphism among six species inNicotiana with two to four alleles (on average 2.61 alleles).
Keyword:Nicotiana; Expressed sequence tags; Resistance gene analogs; SSR Show Figures Show Figures
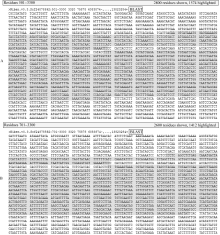
图1 CL1421Contig1、CL1421Contig2与 N. benthamiana基因组序列比对结果A: CL1421Contig1与基因组scaffold (序列ID: Niben.v0.3.Scf24878592)的1078~1448 bp和1547~2749 bp区段匹配;B: CL1421Contig2与基因组支架Scf24878592的1276~1449 bp和1549~2786 bp区段匹配。Fig. 1 Alignment of CL1421Contig1 and CL1421Contig2 to N. benthamianagenomeA: CL1421Contig1 matches with 1078-1448 bp and 1547-2749 bp in genomic scaffold (sequence ID: Niben.v0.3.Scf24878592);B: CL1421Contig2 matches with 1276-1449 bp and 1549-2786 bp in genomic scaffold (sequence ID: Niben.v0.3.Scf24878592).
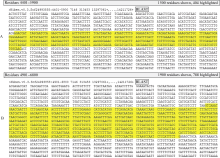
图2 FS423566和CL17188Contig1与烟草基因组序列比对结果A: FS423566与基因组scaffold (序列ID: Niben.v0.3.Scf24993858)的4904~5308 bp区段匹配; B: CL17188Contig1与基因组scaffold (序列ID: Niben.v0.3.Scf24993858)的5494~6282 bp区段匹配。Fig. 2 Alignment of FS423566 and CL17188Contig1 to N. benthamianagenomeA: FS423566 matches with 4904-5308 bp in genomic scaffold (sequence ID: Niben.v0.3.Scf24993858); B: CL17188Contig1 matches with 5494-6282 bp in genomic scaffold (sequence ID: Niben.v0.3.Scf24993858).
EllisJ, DoddsP, PryorT. The generation of plant disease resistance gene specificities. Trends Plant Sci, 2000, 5: 373-379[本文引用:2][JCR: 11.808]
2
SanseverinoW, RomaG, SimoneM D, FainoL, MelitoS, StupkaE, FruscianteL, ErcolanoM R. PRGdb: a bioinformatics platform for plant resistance gene analysis. Nucl Acids Res, 2009, 38: D815[本文引用:2]
3
JohalG S, BriggsS P. Reductase activity encoded by the Hm1 disease resistance gene in maize. Science, 1992, 258: 985-987[本文引用:2]
4
WhithamS, McCormickS, BakerB. The N gene of tobacco confers resistance to Tobacco mosaic virus in transgenic tomato. Proc Natl Acad Sci USA, 1996, 93: 8776-8781[本文引用:1][JCR: 9.737]
5
DixonM S, JonesD A, KeddieJ S, ThomasC M, HarrisonK, Jones J D G, LaneC. The tomato Cf-2 disease resistance locus comprises two functional genes encoding leucine-rich repeat protein. Cell, 1996, 84: 451-459[本文引用:1][JCR: 31.957]
6
BentA F. Plant disease resistance genes: function meets structure. Plant Cell, 1996, 8: 1757-1771[本文引用:1][JCR: 9.251]
7
MeyersB C, DickermanA W, MichelmoreR W, SivaramakrishnanS, SobralB W, YoungN D. Plant disease resistance genes encode members of an ancient and diverse protein family within the nucleotide-binding superfamily. Plant J, 1999, 20: 317-332[本文引用:2][JCR: 6.582]
8
HulbertS H, WebbC A, SmithS M, SunQ. Resistance gene complexes: evolution and utilization. Annu Rev Phytopathol, 2001, 39: 285-312[本文引用:1][JCR: 10.229]
9
DanglJ L, JonesJ D. Plant pathogens and integrated defence responses to infection. Nature, 2001, 411: 826-833[本文引用:1][JCR: 38.597]
10
MeyersB C, KozikA, GriegoA, KuangH, MichelmoreR W. Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell, 2003, 15: 809-834[本文引用:4][JCR: 9.251]
11
BentA F, KunkelB N, DahlbeckD, BrownK L, SchmidtR, GiraudatJ, LeungJ, StaskawiczB J. RPS2 of Arabidopsis thaliana: a leucine-rich repeat class of plant disease resistance genes. Science, 1994, 265: 1856-1860[本文引用:1]
12
DunningF M, SunW, JansenK L, HelftL, BentA F. Identification and mutational analysis of Arabidopsis FLS2 leucine-rich repeat domain residues that contribute to flagellin perception. Plant Cell, 2007, 19: 3297-3313[本文引用:1][JCR: 9.251]
13
SongW Y, WangG L, ChenL L, KimH S, PiL Y, HolstenT, GardnerJ, WangB, ZhaiW X, ZhuL H, FauquetC, RonaldP. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science, 1995, 270: 1804-1806[本文引用:1]
14
TorM, BrownD, CooperA, Woods-TorA, Sjoland erK, JonesJ D, HolubE B. Arabidopsis downy mildew resistance gene RPP27 encodes a receptor-like protein similar to CLAVATA2 and tomato Cf-9. Plant Physiol, 2004, 135: 1100-1112[本文引用:1][JCR: 6.555]
15
MartinG B, BrommonschenkelS H, ChunwongseJ, FraryA, GanalM W, SpiveyR, WuT, EarleE D, TanksleyS D. Map-based cloning of a protein kinase gene conferring disease resistance in tomato. Science, 1993, 262: 1432-1436[本文引用:1]
16
TalerD, GalperinM, BenjaminI, CohenY, KenigsbuchD. Plant eR genes that encode photorespiratory enzymes confer resistance against disease. Plant Cell, 2004, 16: 172-184[本文引用:1][JCR: 9.251]
17
BuschgesR, HollricherK, PanstrugaR, SimonsG, WolterM, FrijtersA, vanDaelen R, van der Lee T, DieRGLardeP, GroenendijkJ, TopschS, VosP, SalaminiF, Schulze-LefertP. The barley Mlo gene: a novel control element of plant pathogen resistance. Cell, 1997, 88: 695-705[本文引用:1][JCR: 31.957]
18
BotellaM A, ColemanM J, HughesD E, NishimuraM T, Jones J D G, SomervilleS C. Map positions of 47 Arabidopsis sequences with sequence similarity to disease resistance genes. Plant J, 1997, 12: 1197-1211[本文引用:1][JCR: 6.582]
19
Aarts M G M, HekkertB L, HolubE B, BeynonJ L, StiekemaW J, PereiraA. Identification of R-gene homologous DNA fragments genetically linked to disease resistance loci in Arabidopsis thaliana. Mol Plant Microbe Interact, 1998, 11: 251-258[本文引用:1][JCR: 4.307]
20
GrahamM A, MarekL F, LohnesD, CreganP, ShoemakerR C. Expression and genome organization of resistance gene analogs in soybean. Genome, 2000, 43: 86-93[本文引用:1][JCR: 1.668]
21
MagoR, NairS, MohanM. Resistance gene analogues from rice: cloning, sequencing and mapping. Theor Appl Genet, 1999, 99: 50-57[本文引用:1][JCR: 3.658]
22
CollinsN C, WebbC A, SeahS, EllisJ G, HulbertS H, PryorA. The isolation and mapping of disease resistance gene analogs in maize. Mol Plant Microbe Interact, 1998, 11: 968-978[本文引用:1][JCR: 4.307]
23
SeahS, SivasithamparamK, KarakousisA, LagudahE S. Cloning and characterization of a family of disease resistance gene analogs from wheat and barley. Theor Appl Genet, 1998, 97: 937-945[本文引用:1][JCR: 3.658]
GaoY L, XuZ L, JiaoF C, YuH Q, XiaoB G, LiY P, LuX P. Cloning, structural features, and expression analysis of resistance gene analogs in tobacco. Mol Biol Rep, 2010, 37: 345-354[本文引用:2][JCR: 2.506]
26
WanH J, ZhaoZ G, MalikA, QianC T, ChenJ F. Identification and characterization of potential NBS-encoding resistance genes and induction kinetics of a putative cand idate gene associated with downy mildew resistance in Cucumis. BMC Plant Biol, 2010, 10: 186[本文引用:1][JCR: 4.354]
27
HuettelB, SantraD, MuehlbauerJ, KahlG. Resistance gene analogues of chickpea (Cicer arietinum L. ): isolation, genetic mapping and association with a Fusarium resistance gene cluster. Theor Appl Genet, 2002, 105: 479-490[本文引用:1][JCR: 3.658]
28
NairR A, ThomasG. Isolation, characterization and expression studies of resistance gene cand idates (RGCs) from Zingiber spp. Theor Appl Genet, 2007, 116: 123-134[本文引用:1][JCR: 3.658]
29
BertioliD J, Leal-BertioliS C, LionM B, SantosV L, PappasG, CannonS B, GuimaraesP M. A large scale analysis of resistance gene homologues in Arachis. Mol Gen Genomics, 2003, 270: 35-45[本文引用:1]
30
MonsiB, WisserR J, PennillL, HulbertS H. Full-genome analysis of resistance gene homologues in rice. Theor Appl Genet, 2004, 109: 1434-1447[本文引用:1][JCR: 3.658]
31
Ameline-TorregrosaC, WangB B, O’BlenessM S, Deshpand eS, ZhuH Y, RoeB, YoungN D, CannonS B. Identification and characterization of nucleotide-binding site-leucine-rich repeat genes in the model plant Medicago truncatula. Plant Physiol, 2008, 146: 5-21[本文引用:2][JCR: 6.555]
32
LiX Y, ChengY, MaW, ZhaoY, JiangH Y, ZhangM. Identification and characterization of NBS-encoding disease resistance genes in Lotus japonicus. Plant Syst Evol, 2010, 289: 101-110[本文引用:4][JCR: 1.312]
33
LiuZ, CramptonM, ToddA, KalavacharlaV. Identification of expressed resistance gene-like sequences by data mining in 454-derived transcriptomic sequences of common bean (Phaseolus vulgaris L. ). BMC Plant Biol, 2012, 12: 42[本文引用:1][JCR: 4.354]
SanzM J, LoarceY, FominayaA, VossenJ H, FerrerE. Identification of RFLP and NBS/PK profiling markers for disease resistance loci in genetic maps of oats. Theor Appl Genet, 2013, 126: 203-218[本文引用:2][JCR: 3.658]
36
LoarceY, SanzM J, IrigoyenM L, FominayaA, FerrerE. Mapping of STS markers obtained from oat resistance gene analog sequences. Genome, 2009, 52: 608-619[本文引用:1][JCR: 1.668]
37
TantasawatP A, PoolsawatO, PrajongjaiT, ChaowisetW, TharapreuksapongA. Association of RGA-SSCP markers with resistance to downy mildew and anthracnose in grapevines. Genet Mol Res, 2012, 11: 1799-1809[本文引用:1][JCR: 0.994]
38
PalominoC, Fernández-RomeroM D, RubioJ, TorresA, MorenoM T, MillánT. Integration of new CAPS and dCAPS-RGA markers into a composite chickpea genetic map and their association with disease resistance. Theor Appl Genet, 2009, 118: 671-682[本文引用:1][JCR: 3.658]
39
FlorH H. The complementary genic systems in flax and flax rust. Adv Genet, 1956, 8: 29-54[本文引用:1][JCR: 4.85]
40
DilbirligiM, GillK S. Identification and analysis of expressed resistance gene sequences in wheat. Plant Mol Biol, 2003, 53: 771-787[本文引用:3][JCR: 3.518]
41
XiaoW K, XuM L, ZhaoJ R, WangF G, LiJ S, DaiJ R. Genome-wide isolation of resistance gene analogs in maize (Zea mays L. ). Theor Appl Genet, 2006, 113: 63-72[本文引用:1][JCR: 3.658]
42
RossiM, AraujoP G, PauletF, GarsmeurO, DiasV M, ChenH, Van Sluys M A, D’HontA. Genomic distribution and characterization of EST-derived resistance gene analogs (RGAs) in sugarcane. Mol Genet Genomics, 2003, 269: 406-419[本文引用:1][JCR: 2.881]
43
XiaoW K, ZhaoJ, FanS C, LiL, DaiJ R, XuM L. Mapping of genome-wide resistance gene analogs (RGAs) in maize (Zea mays L. ). Theor Appl Genet, 2007, 115: 501-508[本文引用:1][JCR: 3.658]
44
HeL, DuC, CovaledaL, XuZ, RobinsonA F, YuJ Z, KohelR J, ZhangH B. Cloning, characterization, and evolution of the NBS-LRR-encoding resistance gene analogue family in polyploid cotton (Gossypium hirsutum L. ). Mol Plant Microbe Interact, 2004, 17: 1234-1241[本文引用:1][JCR: 4.307]
45
PeñuelaS, DaneshD, YoungN D. Targeted isolation, sequence analysis, and physical mapping of nonTIR NBS-LRR genes in soybean. Theor Appl Genet, 2002, 104: 261-272[本文引用:1][JCR: 3.658]
46
BertioliD J, MoretzsohnM C, MadsenL H, Sand alN, Leal-BertioliS C, GuimaraesP M, HougaardB K, FredslundJ, SchauserL, NielsenA M, SatoS, TabataS, CannonS B, StougaardJ. An analyses of synteny of Arachis with Lotus and Medicago sheds new light on the structure, stability and evolution of legume genomes. BMC Genomics, 2009, 10: 45[本文引用:1][JCR: 4.397]
47
BindlerG, PlieskeJ, BakaherN, GunduzI, IvanovN, Van der Hoeven R, GanalM, DoniniP. A high density genetic map of tobacco (Nicotiana tabacum L. ) obtained from large scale microsatellite marker development. Theor Appl Genet, 2011, 23: 219-230[本文引用:1][JCR: 3.658]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}