关键词:普通小麦; puroindoline b-2基因; 产量相关性状; 旗叶大小; 基因定位 Analysis of Association ofpuroindoline b-2 Alleles with Yield-Related Traits in Bread Wheat CHEN Feng, LI Xiang-Nan, CAO Ying-Ying, SUN Jian-Xi, ZHANG Fu-Yan, DONG Zhong-Dong, CUI Dang-Qun* Agronomy College / Collaborative Innovation Center of Henan Grain Crops, Henan Agricultural University, Zhengzhou 450002, China
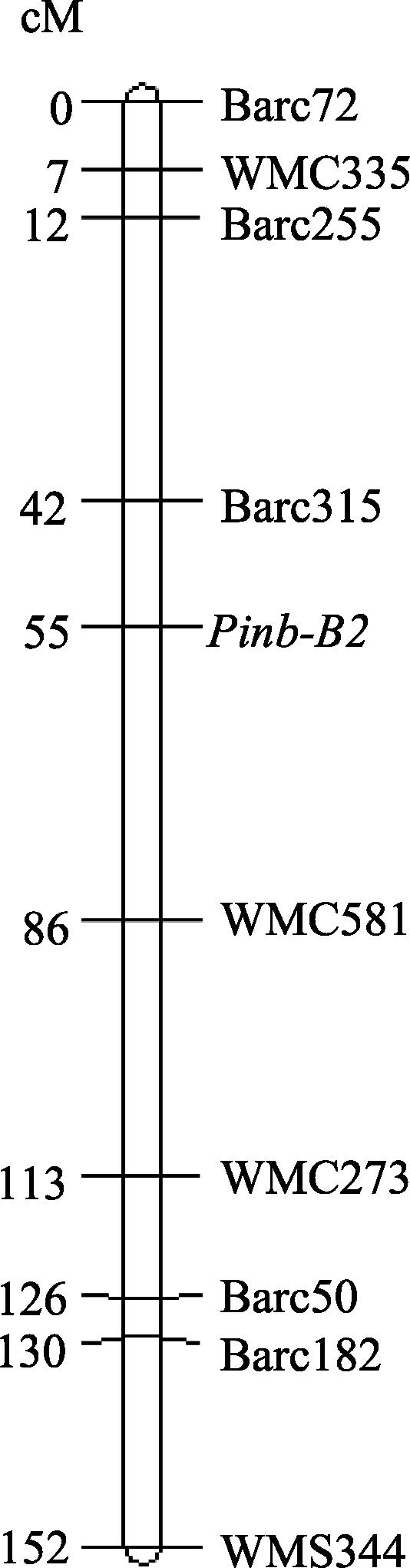
AbstractGenespuroindoline b-2 (Pinb-2) were recently discovered and possess high similarity withpuroindoline b gene. We have identified severalPinb-2 alleles in B genome of bread wheat (Triticum aestivum L.) and found association between allelic variation onPinb-2 and grain texture in soft wheat. In this study, we analyzed the association between thePinb-2 alleles and yield-related traits and flag leaf size, and mapped thePinb-2 locus using a recombinant inbred line (F8) population. Among the fourPinb-2 genotypes,Pinb-2v3b genotype possessed the highest 1000-grain weight, grain diameter, grain number per spike, grain weight per spike, and grain weight per plant. Therefore,Pinb-2v3b genotype possibly has more preferable yield-related traits thanPinb-2v2,Pinb-2v3a, andPinb-2v3c genotypes. Additionally,Pinb-2v3a genotype possessed the highest flag leaf width, length, and area. Genetic mapping indicated thatPinb-2 was located on the long arm of chromosome 7B close to SSR markerBarc315. This result provides basic information for use ofpuroindoline b-2 genes in molecular genetics research and breeding.
Keyword:Bread wheat; puroindoline b-2 genes; Yield-related traits; Flag leaf size; Genetic mapping Show Figures Show Figures
MorrisC F. Puroindolines: the molecular genetic basis of wheat grain hardness. Plant Mol Biol, 2002, 48: 633-647[本文引用:2][JCR: 4.15]
[2]
BhaveM, MorrisC F. Molecular genetics of puroindolines and related genes: allelic diversity in wheat and other grasses. Plant Mol Biol, 2008, 66: 205-219[本文引用:2][JCR: 4.15]
[3]
BhaveM, MorrisC F. Molecular genetics of puroindolines and related genes: regulation of expression, membrane binding properties and applications. Plant Mol Biol, 2008, 66: 221-231[本文引用:3][JCR: 4.15]
[4]
MorrisC F, BhaveM. Reconciliation of D-genome puroindoline allele designations with current DNA sequence data. J Cereal Sci, 2008, 48: 277-287[本文引用:3][JCR: 2.073]
[5]
ChenF, HeZ H, XiaX C, XiaL Q, ZhangX Y, LillemoM, MorrisC F. Molecular and biochemical characterization of puroindoline a and b alleles in Chinese land races and historical cultivars. Theor Appl Genet, 2006, 112: 400-409[本文引用:1][JCR: 3.297]
[6]
ChenF, ZhangF Y, XiaX C, DongZ D, CuiD Q. Distribution of puroindoline alleles in bread wheat of the Yellow and Huai Valley of China and discovery of a novel puroindoline a allele without PINA protein. Mol Breed, 2012, 29: 371-378[本文引用:1][JCR: 2.852]
[7]
陈锋, 董中东, 程西永, 詹克慧, 许海霞, 崔党群. 小麦puroindoline及其相关基因分子遗传基础研究进展. 中国农业科学, 2010, 43: 1108-1116ChenF, DongZ D, ChengX Y, ZhanK H, XuH X, CuiD Q. Advances in research of molecular genetics of puroindoline and its related genes in wheat. Sci Agric Sin, 2010, 43: 1108-1116 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4522]
[8]
MaD Y, ZhangY, XiaX C, MorrisC F, HeZ H. Milling and Chinese raw white noodle qualities of common wheat near- isogenic lines differing in puroindoline b allele. J Cereal Sci, 2009, 50: 126-130[本文引用:1][JCR: 2.073]
[9]
ChenF, LiH H, LiX N, DongZ D, ZuoA H, ShangX L, CuiD Q. Alveograph and Mixolab parameters associated with puroindoline-D1 genes in Chinese winter wheats. J Sci Food Agric, 2013, 10: 2541-2548[本文引用:1][JCR: 1.436]
[10]
ChenF, HeZ H, ChenD S, ZhangC L, XiaX C. Influence of puroindoline allele on milling, steamed bread, noodles and pan bread in common spring wheat. J Cereal Sci, 2007, 45: 59-66[本文引用:1][JCR: 2.073]
[11]
ZhangJ, MartinJ M, Balint-KurtiP, HuangL, GirouxM J. The wheat puroindoline genes confer fungal resistance in transgenic corn. J Phytopath, 2011, 159: 188-190[本文引用:1]
[12]
WilkinsonM, WanY, TosiP, LeveringtonM, SnapeJ, MitchelR A C, ShewryP R. Identification and genetic mapping of variant forms of puroindoline b expressed in developing wheat grain. J Cereal Sci, 2008, 48: 722-728[本文引用:4][JCR: 2.073]
[13]
ChenF, BeecherB, MorrisC F. Physical mapping and a new variant of Puroindoline b-2 genes in wheat. Theor Appl Genet, 2010, 120: 745-751[本文引用:6][JCR: 3.297]
[14]
GengH W, BeecherB S, HeZ H, MorrisC F. Physical mapping of puroindoline b-2 genes in wheat using ‘Chinese Spring’ chromosome group 7 deletion lines. Crop Sci, 2012, 52: 2674-2678[本文引用:5][JCR: 1.641]
[15]
GengH W, BeecherB S, PumphreyM, HeZ H, MorrisC F. Segregation analysis indicates that puroindoline b-2 variants 2 and 3 are allelic in Triticum aestivum and that a revision to puroindoline b-2 gene symbolization is indicated. J Cereal Sci, 2013, 57: 61-66[本文引用:1][JCR: 2.073]
[16]
ChenF, XuH X, ZhangF Y, XiaX C, HeZ H, WangD W, DongZ D, ZhanK H, ChengX Y, CuiD Q. Physical mapping of puroindoline b-2 genes and molecular characterization of a novel variant in durum wheat (Triticum turgidum L. ). Mol Breed, 2011, 28: 153-161[本文引用:5][JCR: 2.852]
[17]
ChenF, LiH H, ShangX L, CuiD Q. A novel puroindoline b-2 variant present in Chinese winter wheat cultivar Yunong 202. J Cereal Sci, 2013, 57: 249-252[本文引用:1][JCR: 2.073]
[18]
RamalingamA, PalomboE A, BhaveM. The Pinb-2 genes in wheat comprise a multigene family with great sequence diversity and important variants. J Cereal Sci, 2012, 56: 171-180[本文引用:4][JCR: 2.073]
[19]
ChenF, ZhangF Y, ChengX Y, MorrisC F, XuH X, DongZ D, ZhanK H, CuiD Q. Association of puroindoline b-B2 variants with grain traits, yield components and flag leaf size in bread wheat (Triticum aestivum L. ) varieties of the Yellow and Huai Valleys of China. J Cereal Sci, 2010, 52: 247-253[本文引用:1][JCR: 2.073]
[20]
张福彦, 陈锋, 董中东, 尚晓丽, 崔党群, 殷贵鸿. 黄淮麦区小麦新品系籽粒硬度相关基因分子鉴定及其对产量相关性状的影响. 中国农业科学, 2011, 44: 3289-3296ZhangF Y, ChenF, DongZ D, ShangX L, CuiD Q, YinG H. Molecular identification of hardness-related genes of bread wheat new lines in Huanghuai wheat region and their influence on yield characters. Sci Agric Sin, 2011, 44: 3289-3296 (in Chinese with English abstract)[本文引用:2][CJCR: 1.4522]
[21]
ChenF, ZhangF Y, DongZ D, MorrisC F, CaoY Y, ShangX L, CuiD Q. Allelic variation of puroindoline b-B2 variant and their independent heredity in bread and durum wheat as well as their association with grain texture. Mol Breed, 2013, 32: 399-409[本文引用:9][JCR: 2.852]
[22]
GengH W, BeecherB S, HeZ H, KiszonasA M, MorrisC F. Prevalence of puroindoline D1 and puroindoline b-2 variants in U. S. Pacific Northwest wheat breeding germplasm pools, and their association with kernel texture. Theor Appl Genet, 2012, 124: 1259-1269[本文引用:1][JCR: 3.297]
[23]
MohlerV, SchmolkeM, PaladeyE, SelingS, HartL. Association analysis of puroindoline-D1 and puroindoline b-2 loci with 13 quality traits in European winter wheat (Triticum aestivum L. ). J Cereal Sci, 2012, 56: 623-628[本文引用:1][JCR: 2.073]
[24]
ZhangJ, MartinJ M, BeecherB, LuC, HannahL C, WallM L, AltosaarI, GirouxM J. The ectopic expression of the wheat puroindoline genes increases germ size and seed oil content in transgenic corn. Plant Mol Biol, 2010, 74: 353-365[本文引用:1][JCR: 4.15]
[25]
GollanP, SmithK, BhaveM. Gsp-1 genes comprise a multigene family in wheat that exhibits a unique combination of sequence diversity yet conservation. J Cereal Sci, 2007, 45: 184-198[本文引用:1][JCR: 2.073]
[26]
ChenF, ZhangF Y, MorrisC F, CuiD Q. Chapter 15: a puroindoline mutigene family exhibits sequence diversity in wheat and is associated with yield-related traits. In: Gene Duplication / Book 2. 2011, pp279-288[本文引用:1]
 , 李向楠, 曹莹莹, 孙建喜, 张福彦, 董中东, 崔党群
, 李向楠, 曹莹莹, 孙建喜, 张福彦, 董中东, 崔党群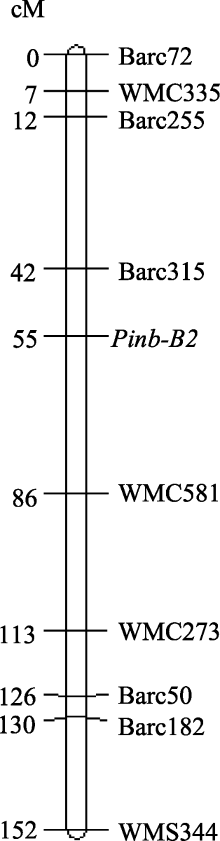

{kind=link}