 , 邓元宝, 李平
, 邓元宝, 李平* 通讯作者(Corresponding authors): 邓其明, E-mail:dengqmsc@163.com, Tel: 1388065608; 李平, E-mail:liping6575@163.com, Tel: 13908070452
第一作者联系方式: E-mail:libin15999211540@sina.com, Tel: 18215636447
收稿日期:2013-06-16 基金:本研究由国家高技术研究发展计划(863计划)项目(2011AA10A101)资助。
摘要7001S是一个广谱抗稻瘟病的粳稻两用核不育系, 对来自全国不同稻区的22株稻瘟病菌系均表现为高度抗性。通过构建7001S/80-4B F2群体的遗传分析和初步定位表明, F2分离单株对稻瘟病菌的抗性呈明显的抗、感双峰分布, 抗感分离符合3﹕1的理论比例, 说明粳稻7001S对稻瘟病菌的抗性由1对显性核基因或一个显性QTL位点控制, 并将该基因初步定位于第11染色体长臂末端。进一步通过扩大遗传群体和分子标记开发, 利用基于BSA的隐性群体分析技术, 将目的基因精细定位于P21-2415和RM27322之间约310 kb的范围内, 并获得了可用于分子标记辅助选择的紧密连锁和共分离分子标记, 同时对目标基因所在区域进行基因预测, 初步确定了候选基因。为进一步开展该抗稻瘟病基因的克隆、功能验证和抗病机理研究, 以及通过分子标记辅助选择技术培育抗稻瘟病水稻新品种等工作奠定了基础。
关键词:稻瘟病; 7001S; 抗性基因; 分子标记; 遗传分析; 精细定位
Identification, Genetic Analysis and Gene Mapping of a Rice Blast Resistance Gene in
LI Bin
, DENG Yuan-Bao, LI PingAbstract7001S is a male-sterile rice with broad-spectrum resistance to rice blast pathogens and highly resistant to 22 strains of
Keyword:Rice blast; 7001S; Resistance gene; Molecular markers; Genetic analysis; Fine mapping
Show Figures
Show Figures




水稻是重要的粮食作物, 世界上有一半以上的人口以稻米为主食。稻瘟病是水稻最严重的病害之一, 分布广泛, 危害严重, 流行年份可造成水稻减产10%~20%, 严重时可减产40%~50%以上, 甚至绝收。据测算, 每年由稻瘟病造成的水稻损失足以养活6000万人以上[ 1]。另外稻瘟病菌不仅侵染水稻, 还能侵染小麦、大麦和粟等禾本科农作物。防治稻瘟病的方法主要为药剂防治和推广抗病新品种。尽管药剂防治对稳定水稻产量起了非常重要的作用, 但所带来的问题也日渐突出: 一方面增加了农民的劳动成本, 另一方面还会造成严重的环境污染, 危及自然界的生物多样性; 同时大量的农药残留对人类的健康构成潜在威胁。实践证明, 利用水稻自身携带的抗病基因是控制稻瘟病害最经济、有效、环保的方法[ 2]。
到目前为止, 在水稻中至少报道了64个抗稻瘟病位点共78个主效基因。这些基因成簇地分布于除第3染色体外的所有水稻染色体上, 其中 Pb1、 Pia、 Pib、 Pi-d2、 Pi-d3、 Pik、 Pik-h/Pi-54、 Pik-m、 Pik-p、 Pi-sh、 Pit、 Pi-ta、 Piz-t、 Pi1、 Pi2、 Pi5、 Pi9、 pi-21、 Pi-25、 Pi-36、 Pi-37、 Pi-50、 Pi-56等23个基因已被成功克隆( Pi2、 Pi9、 Piz-t和 Pi-50同为 Piz位点上的复等位基因; Pi1、 Pik-h/Pi-54、Pik-m、 Pik-p同为 Pik位点上的复等位基因; Pi-d3与 Pi-25等位)(国家水稻数据中心)。在这些已报道的稻瘟病抗性基因中, 除 Pi1、 Pi2、 Pi9、 Pi-20、 Pi-33、 Pi-40、 Pi-gm、 Pik-h等具有广谱抗性外, 其余多数表现为生理小种特异性抗性。同时, 目前所定位或克隆的稻瘟病抗性基因多来源于地方品种或野生稻资源, 而从具有优良农艺性状的育种材料中定位或克隆抗性基因的报道不多见。由于携带这些基因的种质资源本身农艺性状较差, 及在育种实践中不利基因的连锁累赘, 导致这些抗性基因难以被直接利用, 难以满足不同生态区域抗稻瘟病育种目标的要求。因此, 从农艺性状优良的品种中发掘新的广谱抗稻瘟病基因, 将有利于拓宽抗病基因的应用生态区域, 并缩短抗病品种的育种周期从而提高育种效率。
7001S是光敏感核不育水稻农垦58S与常规品系917杂交选育而成的粳型光敏核不育系。经多年在安徽、四川、云南、贵州等稻瘟病重灾区种植鉴定表明, 对稻瘟病表现高度抗性, 是一份理想的稻瘟病抗性研究材料。本研究利用来自全国不同稻区的22株稻瘟病菌系对7001S所含抗性基因系统评价、遗传分析和基因初步定位和精细定位, 获得了可用于分子标记辅助选择的紧密连锁和共分离的分子标记, 同时在对该抗病基因精细定位的基础之上, 对目标基因所在区域进行基因预测, 确定了候选基因。为进一步开展该抗稻瘟病基因的克隆、功能验证和抗病机制研究, 以及通过分子标记辅助选择技术培育抗稻瘟病水稻新品种等方面的工作奠定了基础。
1 材料与方法1.1 植物材料抗性基因供体材料为粳稻两系不育系7001S (安徽省农业科学院水稻研究所提供)。高感稻瘟病材料为粳稻80-4B (安徽省农业科学院水稻研究所提供)。诱发品种为江南香糯(四川农业大学水稻研究所保存)。
1.2 供试菌系用于稻瘟病抗性鉴定的稻瘟病菌系ZA13、ZB3、ZB9、ZB13、ZB14、ZB15、ZC15、ZD7、ZF1和ZG1由本实验室引进保存; 北1(007)、ZH2-1、91-13-2、91-17-2和97-65-2由中国农业科学院作物科学研究所稻病室提供; 菌株11-14、11-15、95-51-1、95-53-1、95-56-1、97-28-1和97-41-1由四川农业大学水稻研究所分别从芦山县眉东镇和雅安市草坝镇病株活体采样分离获得。
1.3 亲本抗性鉴定、抗谱分析和强致病菌株筛选2011年夏季在温江和雅安草坝将抗病品种7001S、感病品种80-4B、7001S/80-4B F1和诱发品种江南香糯按22株接种菌株分组隔离种植。4月8日播种, 5月12日移栽。翻耙时按750 kg hm-2施水稻专用复合肥作底肥。分别在三叶期和分蘖期追施尿素90 kg hm-2, 促进秧苗生长嫩绿, 利于病害发生和流行, 实验过程中采用生物防治法防范其他病虫害。分别于移栽后2周和1个月用鉴定菌菌株的孢子悬浮液喷雾接种, 接种后覆膜保湿24 h促进发病。7月下旬, 在叶瘟完全稳定后调查病情并记录, 并筛选出强致病菌株用于遗传分析群体接种。叶瘟以最高级为鉴定结果, 按国际水稻研究所9级制标准调查记录, 0~3级为抗病, 5~9级为感病。抗性分级标准如下:0级: 无病斑; 1级: 叶片上产生针头状大小的褐点型病斑; 2级: 稍大病斑; 3级: 小圆形稍长的灰色病斑, 边缘褐色, 病斑直径1~2 mm; 4级: 典型的纺锤形病斑, 长1~2 cm, 通常局限在两条主脉间, 危害面积不超过叶面积的2%; 5级: 典型病斑, 危害面积不超过叶面积的10%; 6级: 典型病斑, 危害面积为叶面积的11%~25%; 7级: 典型病斑, 危害面积在叶面积的26%~50%; 8级: 典型病斑, 危害面积为叶面积的51%~75%; 9级: 典型病斑, 危害面积为叶面积的76%至全叶枯死。
1.4 遗传群体构建2011年夏季在雅安草坝种植7001S/80-4B F2群体。4月8日播种, 5月12日移栽, 单苗移栽, 栽插规格为17 cm × 23 cm, 翻耙时按750 kg hm-2施水稻专用复合肥作底肥。然后分别在三叶期和分蘖期追施尿素90 kg hm-2, 促进秧苗生长嫩绿, 利于病害发生和流行, 实验过程中采用生物防治法防范其他病虫害。分别于移栽后15 d和30 d用筛选出的强致病菌株孢子悬浮液喷雾接种, 接种后覆膜保湿24 h促进发病。7月下旬, 在叶瘟完全稳定后调查病情并记录。
1.5 DNA提取、PCR扩增及电泳分析在接种结果调查完后, 分别取抗病亲本7001S、感病亲本80-4B及7001S/80-4B的F2群体极端感病单株和抗性单株的叶片提取DNA。采用高效便捷的水稻叶片DNA提取方法[ 3], 将其溶解在TE缓冲液里(10 mmol L-1 Tris base, 0.1 mmol L-1 EDTA)。每份DNA统一用ddH2O稀释成20 ng μL-1, 作为PCR扩增分析的模板。用本实验室已有的均匀分布于水稻12条染色体上的972对SSR引物直接在抗、感亲本间筛选多态性标记。以上引物由上海英潍捷基贸易有限公司合成。20 μL的PCR体系含 DNA模板2 μL (<0.5 μg)、SSR引物2 μL (0.2~0.4 μmol L-1)、10×buffer (含Mg2+) 2 μL、dNTP (2.5 mmol L-1) 0.5 μL、 Taq DNA聚合酶0.2 μL (2.5 U)、ddH2O 13.3 μL。在美国Thermo公司PCR1000上进行扩增反应, 94℃预变性5 min, 94℃变性45 s、55℃退火45 s、72℃延伸1 min、32个循环, 72℃延伸10 min。扩增产物经3%的琼脂糖凝胶电泳直接检测。
1.6 BSA与RCA分析以粳稻抗病品种7001S与普感稻瘟病品种80-4B杂交得到F2群体作为精细定位群体, 含376个极端感病单株和50个抗病单株。采用F2分离群体分池法(BSA)结合隐性类型分析法(RCA), 对分子标记和抗性基因进行连锁遗传分析。具体步骤如下: (1)分别提取抗病亲本7001S、感病亲本80-4B和江南香糯、以及F2代分离群体中部分抗病植株和全部感病单株的DNA, 选取10个抗病株DNA等量混合, 构建抗病池(R池); 选取10个感病株DNA等量混合, 构建感病池(S池)。(2)利用分子标记对抗病亲本和感病亲本进行多态性分析, 筛选出具有多态性的标记; 然后用在抗感亲本间具有多态性的标记对抗、感病池DNA进行扩增, 获得在抗、感病池间有多态性的标记。(3)利用池间多态性标记对由100株极端感病单株(隐性个体)组成的小群体进行PCR检测, 进一步确定是否存在连锁关系。如果连锁, 用以对扩大隐性单株群体进行扩增和连锁分析, 并用Mapmaker/exp 3.0软件中的Mapdraw构建更加紧密的分子连锁图。
1.7 遗传图谱的构建当目标基因被两侧的标记界定之后, 再用两侧最近的标记分析F2群体, 再用新开发的标记对所有重组体进行检测, 根据重组体的减少情况逐步将目的基因定位到更小区域内, 根据所有标记与目标基因之间的遗传距离整合成一张遗传图。再利用BLASTN分析, 将各个与目标基因连锁的标记锚定到物理重叠群上, 标明各标记的物理位置和在各自位点发生的重组数以及各个标记之间的物理距离。
2 结果与分析2.1 7001S稻瘟病抗谱分析将抗病品种7001S、感病品种80-4B、7001S/80-4B F1和诱发品种江南香糯按接种菌株分组隔离种植。每组均种植抗病品种7001S及感病品种80-4B各2行, 每行10株, 用江南香糯诱发。接菌鉴定方法同抗病分析中采用的方法, 待叶瘟完全稳定后调查病情, 叶瘟以最高级为鉴定结果, 并按国际水稻研究所9级制记录。
从表1可以看出, 在温江和雅安两地, 7001S对22个鉴定菌株均表现出高度抗性, 抗级反应为0~3级, 而感病品种80-4B对所有鉴定菌系均表现感病, 其抗级反应为5~9级。所有菌株均能较充分地发病, 其中ZB13、ZB15、97-28-1和97-41-1致病力最强, 发病最充分。由此推断7001S对稻瘟病抗性较强, 抗性稳定, 且抗谱广, 是一份具有重大应用前景的粳稻抗性材料, 预示其可能含有重要的稻瘟病抗性基因或稻瘟病抗性QTL存在, 具有进一步深入研究和开发的潜力。并将强致病标准菌株ZB15确定为本研究后续研究接种菌株。
表1
Table 1
表1(Table 1)
| 表1 亲本抗性鉴定调查结果 Table 1 Result of parents’ resistance identification | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2.2 7001S/80-4B杂种F1代的抗性表现 为了进一步明确7001S所含稻瘟病抗性基因的真实性及其基因的显/隐性关系, 以亲本抗性鉴定和抗谱分析中发病最充分的菌株ZB15对7001S/80-4B F1接种鉴定。从表2可以看出, 7001S/80-4B杂种F1代无论对叶瘟还是穗颈瘟均表现为高抗, 说明7001S所含目标抗性基因或抗性QTL表现为显性, 预示其在粳稻杂交水稻育种上具有广泛的应用前景。
表2
Table 2
表2(Table 2)
| 表2 7001S/80-4B杂种F1代的抗性鉴定结果 Table 2 Resistance identification in F1 population of cross between 7001S and 80-4B | |||||||||||||||||||||||||||||||||||
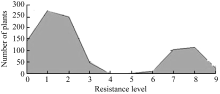
2.3 抗性位点的遗传分析F2单株群体人工接种稻瘟病菌后, 待叶瘟完全稳定后(图1)调查病情, 调查及记录各株水稻的发病情况和病斑的大小和多少, 以最高级为鉴定结果, 调查记载标准按国际水稻研究所9级制进行抗感分级。将统计结果绘制成分布曲线图(图2)。从图2可以看出, F2单株对稻瘟病的抗性呈明显的抗、感分布, 可清晰地划分为抗病集团和感病集团。分别将分布于抗性集团和感病集团的单株数统计和卡方检验, 结果表明: 在F2代群体的946株中, 抗稻瘟病单株为694株, 感病植株252株, 分离比为2.75∶1.00, 卡方检验χ2 = 1.3545 (χ20.05,1 = 3.84), 符合3∶1的理论比例, 说明7001S对稻瘟病菌的抗性由1对显性核基因或一个显性QTL位点控制。
图1
Fig. 1
| Figure OptionViewDownloadNew Window | |
 | 图1 稻瘟病抗病区与感病区的对照图Fig. 1 Cross reference map of the resistant area and susceptible area to the blast |
图2
Fig. 2
| Figure OptionViewDownloadNew Window | |
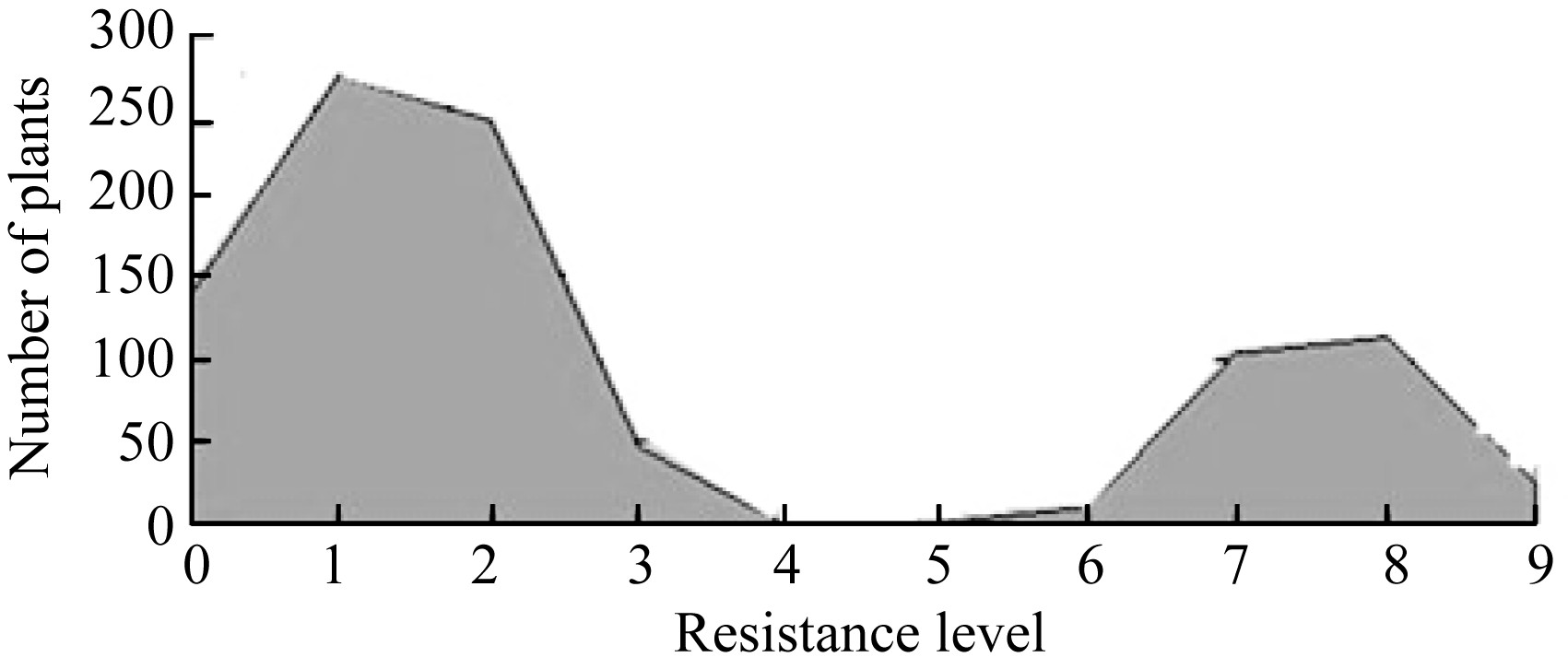 | 图2 F2群体抗感分布0~3抗病株; 6~9感病株。 0-3: resistant plants; 6-9: susceptible plants.Fig. 2 Distribution of the resistance and susceptibility in F2 population |
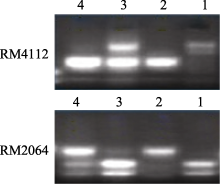
2.4 多态性标记筛选和抗性基因初步定位用本实验室已有的均匀分布于水稻12条染色体上的972对SSR引物对抗、感亲本进行多态性分析, 获得多态性标记78对。然后分别利用F2群体中的感病和抗病单株建立抗病池和感病池, 对78对多态性标记进一步筛选, 获得抗病池和感病池间的多态性标记RM4112和RM2064 (图3)。利用RM4112和RM2064对由100株极端感病单株(隐性个体)组成的小群体进行PCR检测表明, 目标基因与其具紧密连锁关系, 目标基因被初步定位于第11染色体长臂末端RM4112和RM2064之间, 并将该抗稻瘟病基因暂时命名为 Pi-ja。
图3
Fig. 3
| Figure OptionViewDownloadNew Window | |
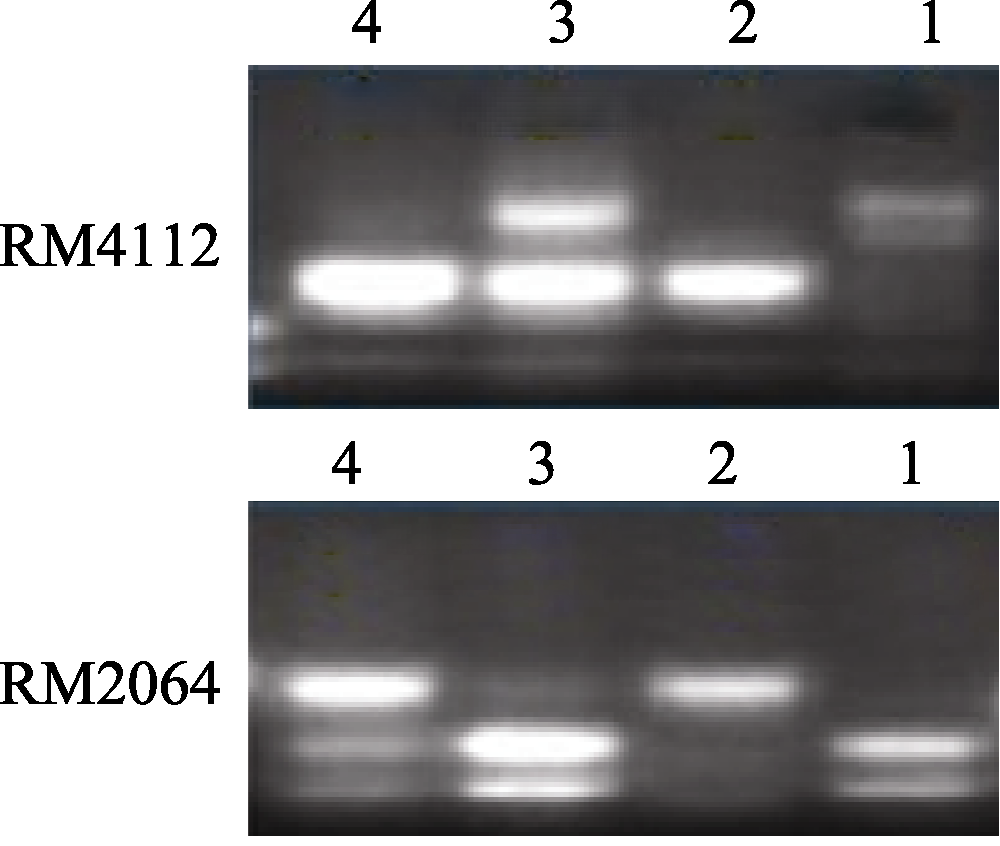 | 图3 SSR标记在池中的电泳分离图1: 7001S; 2: 80-4B; 3: 抗病池; 4: 感病池。Fig. 3 Segregation patterns of the SSR markers in the pool1: 7001S; 2: 80-4B; 3: resistant pool; 4: susceptible pool. |
2.5 抗性基因精细定位在初步定位基础上, 在目标基因区域开发了相应的SSR、InDel和CAPS标记, 其中有9个标记与
目标基因连锁, 分别是S3-13、I7-1、P21-2415、N22-550、N22-3600和S10-4 (表3和图4)。其中S3-13检测到2个重组体, 位于RM27274一侧; P21-2415只检测到了1个重组体也位于RM27274一侧, 而I7-1、N22-550、N22-3600三个标记没有检测到重组体, 说明这3个标记和目标基因共分离。另一个标记S10-4检测到了1个重组体与RM27322检测到的重组体相同, 说明S10-4位于RM27322一侧。最终将目的基因定位在P21-2415和RM27322之间, 且与标记I7-1、N22-550、N22-3600共分离。
表3
Table 3
表3(Table 3)
| 表3 在F2群体中与抗性基因连锁的标记 Table 3 Markers linked to resistance gene in F2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
图4
Fig. 4
| Figure OptionViewDownloadNew Window | |
 | 图4 S3-13、P21-2415、N22-550、N22-3600、S10-4在F2群体中重组体检测P1: 7001S; P2: 80-4B; 1~22: 极端感病个体; S3-13中3、21, P21-2415中3, S10-4中16是重组单株。Fig. 4 Recombinant detection in F2 population by S3-13 , P21-2415, N22-550, N22-3600, and S10-4P1: 7001S; P2: 80-4B; 1-22: susceptible individuals; 3, 21 in S3-13, 3 in P21-2415, 16 in S10-4 are recombinated individuals. |
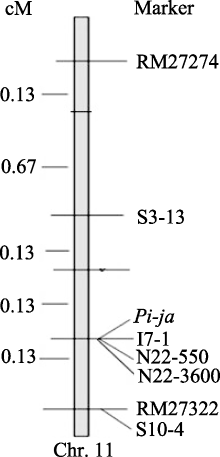
2.6 Pi-ja基因区域精细遗传连锁图 根据上述目标区域分子标记与目标基因 Pi-ja的遗传重组情况, 进一步整合遗传距离, 将连锁的标记整合到遗传图谱上。在F2群体中 Pi-ja被定位在P21-2415和RM27322之间的0.27 cM区域内, 目标基因 Pi-ja与这2个标记之间均只发生了1次重组, 并与I7-1、N22-550、N22-3600三个标记共分离(图5)。
图5
Fig. 5
| Figure OptionViewDownloadNew Window | |
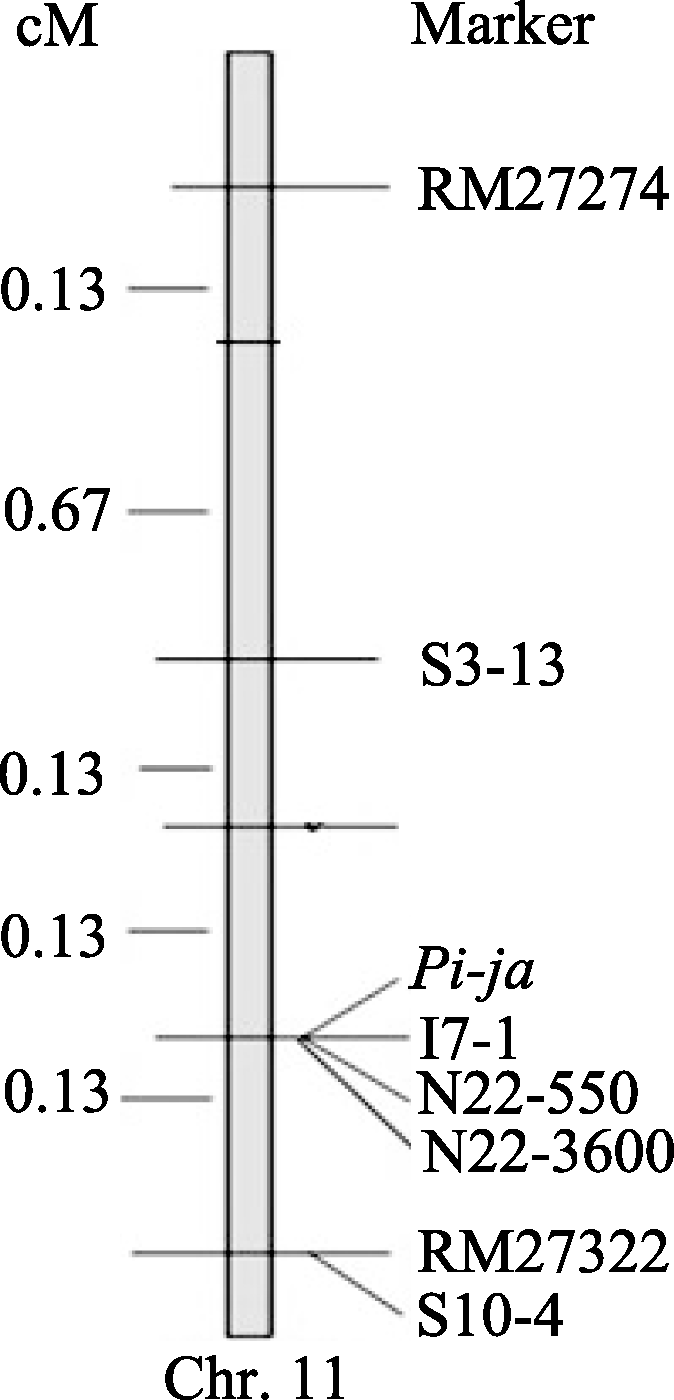 | 图5 F2群体中 Pi-ja遗传连锁图Fig. 5 Genetic linkage map of Pi-ja in F2 |
2.7 Pi-ja基因区域物理图构建 根据各标记在物理重叠群上着陆的位置, 构建了 Pi-ja基因区域的电子物理图(图6)。从RM27274到P21-2415之间一共9个重叠克隆组成的重叠群覆盖了 Pi-ja基因区域, 本研究中用于定位 Pi-ja基因的主要分子标记被锚定在此图上, 目的基因最终被定位在标记P21-2415和RM27322之间约310 kb的范围内。
图6
Fig. 6
| Figure OptionViewDownloadNew Window | |
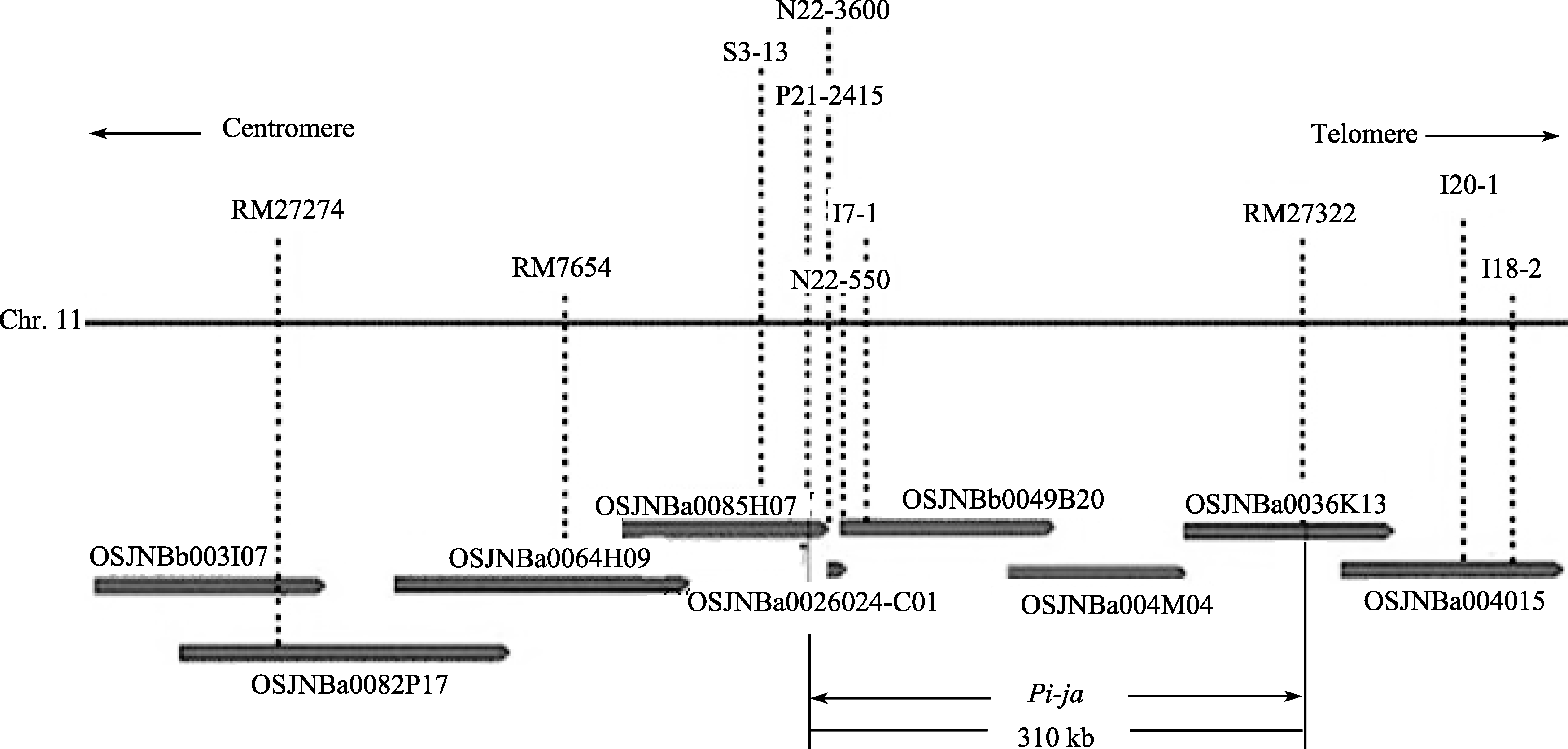 | 图6 Pi-ja基因区域的物理图  Fig. 6 Physical mapping of Pi-ja gene locus Fig. 6 Physical mapping of Pi-ja gene locus  |
2.8 候选基因的预测与分析利用RGP在线基因预测软件RiceGAAS (http:// ricegaas.dna.affrc.go.jp/), 结合在线数据库GRAMENE 、TIGR、NCBI的基因注释, 在 Pi-ja定位区间内确定了3个具有抗病基因结构特征的候选基因(表4), 其中 LOC_Os11g45980和 LOC_Os11g46100两个是NBS- LRR类型的基因, LOC_Os11g46200是LRR类型基因。
表4
Table 4
表4(Table 4)
| 表4 目标区域基因的预测结果 Table 4 Results of gene prediction in target regions |
测序结果表明, 候选基因 LOC_Os11g46100全长1.6 kb, 在抗性亲本7001S和感病亲本80-4B之间序列完全相同, 因此将 LOC_Os11g46100排除。候选基因 LOC_Os11g46200编码LRR类抗病蛋白, 含3个外显子, 基因总长3378 bp, 编码1125个氨基酸, 对该基因的蛋白质结构预测显示, 它包含1个ATP激酶结合位点和一个富亮氨酸重复序列结构域。候选基因 LOC_Os11g45980在Gramene上的注释是编码NBS-LRR抗病蛋白, 含有3个外显子, 基因总长3821 bp, 编码1135个氨基酸。测序发现其基因序列、cDNA和氨基酸序列在亲本间均存在差异: 在抗性亲本7001S中 LOC_Os11g45980具有4个外显子, 基因总长5255 bp, cDNA全长2976 bp, 编码991个氨基酸; 而在感病亲本80-4B中 LOC_ Os11g45980仅具有2个外显子, 基因总长5272 bp, cDNA全长3402 bp, 编码1133个氨基酸(图7)。根据序列差异设计了2个CAPS标记N22-3600和N22-550, 2个标记在定位群体中均未检测到重组体, 表现为共分离。
图7
Fig. 7
| Figure OptionViewDownloadNew Window | |
 | 图7 7001S和80-4B中LOC_Os11g45980氨基酸序列的差异阴影部分表示抗感亲本氨基酸差异。Fig. 7 Difference of LOC_Os11g45980 amino acid sequence in 7001S and 80-4BThe shading parts indicate amino acid differences between resistant and susceptible parents. |
3 讨论近年来, 各国科学家对稻瘟病的发生机理以及水稻的抗性机制进行了大量研究并取得了巨大突破。然而, 由于稻瘟病小种的快速变异以及环境的复杂性, 含有单一抗病基因的品种种植几年后就会失去其原有的抗性。因此, 挖掘新的抗病基因资源, 采用分子标记进行多基因聚合育种, 培育新的具有广谱抗性的水稻品种成为水稻抗病育种的关键。7001S对ZA13、ZB3、ZB9、ZB13、ZB14、ZB15、ZC15、ZD7、ZF1和ZG1, 中国农业科学院作物科学研究所稻病室提供的北1 (007)、ZH2-1、91-13-2、91-17-2和97-65-2, 以及由四川农业大学水稻研究所分别从芦山县眉东镇和雅安市草坝镇病株活体采 样分离获得的11-14、11-15、95-51-1、95-53-1、95-56-1、97-28-1和97-41-1等22株强致病性菌株都有很好的抗性, 是理想的稻瘟病抗性资源。该基因的鉴定、定位克隆和应用, 将对我国水稻特别是粳稻稻瘟病抗性改良发挥重要的作用。
稻瘟病抗性基因遗传相当复杂, 一般由1个或2个主效 R基因控制, 有些由3个或更多主效 R基因控制, 少数情况下由隐性 R基因或不完全显性 R基因控制, 不同 R基因间相互独立或存在互作关系[ 4]。到目前为止在水稻中至少报道了64个抗稻瘟病位点, 其中已经成功克隆24个抗病基因[ 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16], 同时也克隆了7个[ 8]对应的无毒基因。从染色体分布来看, 这些抗性基因在水稻基因组第3染色体以外的其余11条染色体均有分布, 但在第6、第11和第12染色体上分布尤为集中。这些已克隆的抗稻瘟病基因中除 Pi-21[ 17]为隐性抗病基因外, 均是显性基因; 除 Pi-21和 Pb1[ 18]为数量性状基因外, 都是质量性状的抗性基因; 从基因结构特征看, 除 Pi-21和 Pi-d2[ 19]分别编码一个富含脯氨酸蛋白和类受体激酶外, 都属于NBS-LRR类基因。另外, 在长臂末端SSR标记RM1233~RM27369之间存在一个大的基因簇[ 20], 包含 Pik[ 21]、 Pik-p[ 22]、 Pik-m[ 5]、 Pi1[ 7]、 Pi5[ 22]和 Pia[ 23]6个基因, 这些基因均需要其对应基因座的两个紧密连锁的NBS-LRR基因同时存在才能发挥抗病性。 Pik位点存在多个等位基因可能是平衡选择的结果[ 21]。平衡选择理论认为自然选择的作用没有使某基因位点的等位基因频率上升或下降, 而是使该基因座上2种或2种以上的等位基因保持平衡[ 24]。Tian等[ 25]发现抗病基因呈等位分布的情况在其他植物中也广泛存在, 比如小麦抗白粉病基因[ 26]位点与亚麻抗锈病基因L位点[ 27], 也发现拟南芥抗病基因座RPS5同时存在多个功能等位基因, 并认为这是植物抵抗病原菌的一种选择机制。本研究采用BSA和RCA分析方法,将7001S的广谱抗性基因 Pi-ja精细定位在第11染色体的标记P21-2415和RM27322间约310 kb的范围内。定位标记等位分析结果表明, pik-h和 pi-54在标记RM224和Y6855A之间2 cM, 而 pi-54和RM206紧密连锁。 Pik-m在RM254和RM144之间1.2 cM, 并且与之紧密连锁, 而 Pi-ja定位于P21-2415和RM27322之间。经Gramene数据库比对发现 Pi-ja在 Pik-m和 Pik的定位标记范围之内, 但与 Pi-54 ( Pik-h)和 Pik-p等位位置略有不同。说明 Pi-ja可能属于 Pik基因簇的一个等位变异基因。但本研究发现的主效抗性基因 Pi-ja与 pik、 Pik-m及 Pik-p的具体关系, 还有待进一步深入研究。
近年全国两系杂交稻种植面积占杂交稻播种面积的25%以上[ 31], 两系法已成为水稻杂种优势利用的主要途径之一。然而, 目前生产上使用的水稻两用核不育系对稻瘟病抗病和耐病能力普遍较低[ 32], 因此培育对稻瘟病具有持久广谱抗性的粳型两用核不育系将对两系杂交水稻的发展具有重要作用。本研究精细定位7001S的主效抗性基因 Pi-ja, 一方面为进一步通过图位克隆该抗性基因奠定了基础; 另一方面, 获得的与目标基因紧密连锁的分子标记P21-2415和RM27322, 为通过分子标记辅助选择技术改良水稻两用核不育系的稻瘟病抗性提供了实用的分子标记。
4 结论来源于粳稻两系不育系7001S的抗稻瘟病基因受1对显性基因控制, 对来自全国不同稻区的稻瘟病菌株均表现较强的抗性, 具有抗性强、抗谱广的特点, 是一份理想的稻瘟病抗性资源。将目的基因定位在第11染色体上P21-2415和RM27322之间约310 kb的范围内, 构建了 Pi-ja基因区域的精细遗传连锁图谱和物理图谱, 开发了与目的基因共分离的分子标记I7-1、N22-550和N22-3600, 此标记已经完全满足分子标记辅助选择的要求, 在目的基因成功克隆之前, 可以利用这些分子标记开展分子标记辅助育种, 将目标基因导入目前大面积推广的粳稻品种中, 改良粳稻品种的稻瘟病抗性。获得2个候选基因 LOC_Os11g45980和 LOC_Os11g46200, 目前正在进行转基因功能验证和干涉实验验证其功能。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] | |
| [25] | |
| [26] | |
| [27] | |
| [28] | |
| [29] | |
| [30] | |
| [31] | |
| [32] |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}