 , 蒋雷, 鞠丽萍, 金秀锋, 王轩, 张晓科, 王宏礼, 付晓洁
, 蒋雷, 鞠丽萍, 金秀锋, 王轩, 张晓科, 王宏礼, 付晓洁* 通讯作者(Corresponding author): 张晓科, E-mail:zhangxiaoke66@126.com
第一作者联系方式: E-mail:zf2606@163.com
收稿日期:2013-08-16 基金:本研究由国家自然科学基金项目(30971770), 引进国际先进农业科学技术计划(948计划)项目(2011G-3)和美国唐仲英基金会资助。
摘要抗旱相关基因的挖掘和分子标记开发对选育抗旱小麦品种有重要意义。采用同源克隆、电子克隆、RACE技术和生物信息学分析手段, 获得了普通小麦硫氧还蛋白(Trx)超家族一个新基因(
关键词:普通小麦; 抗旱性; TaNRX; 分子标记
Cloning a Novel Gene
ZHANG Fan
, JIANG Lei, JU Li-Ping, JIN Xiu-Feng, WANG Xuan, ZHANG Xiao-Ke, WANG Hong-Li, FU Xiao-JieAbstractMolecular markers associated with genes for drought resistance play an important role in wheat breeding aiming at improvement of drought resistance. In this study, we obtained the full-length cDNA of a novel gene of thioredoxin (Trx) superfamily,
Keyword:Common wheat; Drought resistance; TaNRX; Molecular markers
Show Figures
Show Figures




小麦是我国重要的粮食作物, 干旱是限制小麦生产的重要因素[ 1, 2]。选育和推广抗旱小麦品种是
保障国家粮食安全、促进小麦生产持续稳定发展的有效途径之一。抗旱基因的挖掘和分子标记开发,对小麦育种具有重要实践意义。
干旱胁迫会引起植物体内一系列生理生化功能的改变, 导致呼吸增强、光合降低、生物膜结构破坏等[ 3]。应对干旱胁迫, 植物体内诱导许多基因的特异表达, 以维持代谢平衡[ 4]。干旱胁迫下植物体内蛋白质组变化的研究, 有助于寻找抗旱相关的蛋白和基因。Caruso等[ 5]发现干旱胁迫下小麦蛋白质组可鉴定的36个差异表达蛋白点中, 有15%与活性氧清除功能相关, 且均显著上调表达; Peng等[ 6]也指出在受到干旱胁迫时, 抗旱小麦品种可表达较多与抗氧化有关的蛋白。活性氧的积累是导致植物在干旱胁迫下细胞组分损伤的直接原因之一, 植物体内活性氧的清除主要依靠一些抗氧化保护的酶类[ 7]。硫氧还蛋白(thioredoxin, Trx)是一类广泛存在于植物体内的多功能蛋白, 它能参与光合作用中的电子传递、被氧化蛋白质的修复、体内活性氧的清除等生理代谢[ 8, 9, 10]。夏德习等[ 11]研究发现, 拟南芥硫氧还蛋白M1型基因( AtTRXm1)在H2O2的处理下大量表达, 表明其与氧化胁迫相关。Trx与含有二硫巯基的许多蛋白组成一个超家族, Broin等[ 12]发现1个干旱诱导蛋白CDSP32, 包含两个与Trx类似的肽序列结构, 为Trx超家族成员, 其mRNA和蛋白受氧胁迫的诱导而大量表达, 认为该蛋白能缓解干旱引起的氧化胁迫。Laughner等[ 13]在玉米中首次分离出了作物体内的核氧还蛋白(nucleoredoxin, NRX)基因, 其编码产物属于Trx超家族的新成员, 包含3个Trx活性功能区, 但其涉及的代谢途径和功能尚不清楚, 目前在其他作物中未见相关基因的报道。
本实验室在前期研究旱地小麦品种晋麦47幼苗叶片水分胁迫蛋白质组表达差异中, 发现一个显著上调的蛋白质谱, 鉴定为大麦某个预测蛋白(GenBank登录号为BAJ87919), NCBI数据库中暂无小麦与之同源的序列。为进一步明确该水分胁迫差异蛋白信息, 挖掘小麦抗旱相关基因, 本研究采用同源克隆和RACE等技术, 在普通小麦中克隆到一个 TaNRX基因, 并基于两组极端抗旱品种该基因的序列差异开发分子标记, 旨在为抗旱小麦品种选育提供依据。
1 材料与方法1.1 试验材料及萌发期抗旱性鉴定以旱地小麦品种晋麦47为材料进行目的基因cDNA克隆。
按照GB/T21127-2007“小麦抗旱性鉴定评价技术规范”测定各基因型萌发期的相对发芽率(-0.5MPa PEG-6000水溶液培养7d发芽率与无离子水培养7 d发芽率的比值), 并按相对发芽率 ≥90.0%、70.0%~89.9%、50.0%~69.9%、30.0%~ 49.9%、≤29.9%依次划分为极强、强、中等、弱和极弱五种抗旱等级。
1.2 总RNA提取及目的基因cDNA克隆将晋麦47的种子放置于以滤纸为芽床的培养皿内, 在光照培养箱中20℃暗培养至幼苗高20 cm左右, 继续在光照强度300 μmol m-2 s-1条件下培养2 d, 然后用-0.5 MPa的PEG-6000处理48 h, 采集叶片用Trizol试剂盒(北京百泰克生物技术公司)提取总RNA。
以本实验室前期所获得的水分胁迫差异蛋白点的质谱测序结果序列(GenBank登录号为BAJ87919)为探针, 在NCBI网站上通过BLAST对比, 筛选出一致性(Identities) ≥70%和相似性(Positives) ≥ 80%的12个禾本科同源蛋白序列(表1)。在保守区设计RT-PCR引物F1 (5′-ACTGTAGCTATCTATT TCTCG-3′)和R1 (5′-TCTTTGCGTCCGTGTTGA-3′), 扩增程序为94℃ 5 min; 94℃ 30 s, 55℃ 30 s, 72℃ 2 min, 35个循环; 72℃ 10 min。再以测序结果为探针, 在NCBI小麦EST数据库中BLAST, 找到1条EST片段(GenBank登录号为HX179294), 并构建EST跨叠群, 设计特异引物F2 (5′-AACAAACGGCATAAA CCA-3′)和R2 (5′-TCAGGACCAATCAGCACTA-3′), 以胁迫后总RNA反转录产物为模板扩增获得基因的5′端序列片段, 扩增程序同上。然后按3′ RACE Kit(北京百泰克生物技术公司)的使用要求设计嵌套的3′ RACE异性引物F3 (5′-GAATCCCTTCTCTG GTCGCCATTGG-3′)和F4 (5′-AAGACGGTCAACAC GGACGCAAAGA-3′); 以干旱胁迫后总RNA为模板, 依照试剂盒操作说明进行3′RACE, 获得目的基因的3′端序列片段。回收纯化PCR产物, 连接至载体pMD18-T, 转化大肠杆菌DH5α, 鉴定后送南京金斯瑞生物科技公司测序。最后使用DNAMAN软件拼接, 得到全长cDNA序列。
表1
Table 1
表1(Table 1)
| 表1 高相似性候选蛋白的BLAST结果 Table 1 BLAST results of candidate proteins with high similarity |
1.3 序列的生物信息学分析使用NCBI数据库的BLAST程序比对分析同源性, DNAMAN软件查询开放阅读框, 采用SOPMA (http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl)在线软件分析蛋白质的二级结构[ 14]; 利用EXPASY网站
(
1.4 基因结构分析与标记开发采用CTAB法[ 20]提取黄化苗基因组DNA。依据cDNA序列设计3对引物(表2)。在利用中国春的双体、缺体-四体和双端体对引物进行染色体定位的基础上, 以晋麦47和西农2208的基因组DNA为模板分段扩增目标基因的基因组DNA序列。PCR扩增程序为94℃预变性5 min后, 94℃变性30 s, 51、54或55℃退火30 s, 72℃延长1~2 min, 30个循环; 最后72℃延长10 min。回收PCR产物并测序, 所用方法同1.2部分。拼接基因组DNA序列后与cDNA序列进行比较分析。为明确该DNA序列差异与小麦抗旱性之间的关系, 选用晋麦47和西农2208 (水地品种)进行目的基因的基因组序列差异分析, 进一步选择两个相对发芽率高于90%的抗旱性极强品种鲁麦21 (97.30%)和临旱917 (92.00%), 与2个相对发芽率低于30%的抗旱性极弱品种济麦20 (20.90%)和陕150 (26.00%)进行目的基因序列测序比较。依据差异序列设计4对引物(表3)。
以中国春的双体、缺体-四体和双端体材料, 对目的基因及其分子标记作染色体定位。在此基础上, 用来自全国不同地区的150份品种(系)进行标记验证。PCR程序为94℃ 5 min; 94℃ 30 s, 52~57℃ 30 s, 72℃ 1 min, 30个循环; 72℃ 10 min。
表2
Table 2
表2(Table 2)
| 表2 克隆 TaNRX基因引物 Table 2 Primers used for cloning TaNRX gene |
表3
Table 3
表3(Table 3)
| 表3 TaNRX基因分子标记 Table 3 Molecular markers for TaNRXgene |
2 结果与分析2.1 目标基因全长cDNA克隆利用引物F1和R1对晋麦47进行同源克隆, 获得1270 bp的片段。BLAST比对结果发现, 与其同源性最高的为大麦的一个克隆(GenBank登录号为AK356704), 其恰好是用于保守位点分析的蛋白探针(GenBank登录号为BAJ87919)的核苷酸序列, 从而证明该片段为目标基因cDNA片段。通过电子克隆得到971 bp含起始密码的5′端序列, 包含752 bp的重叠区; 3′RACE获得长度为551 bp的3′端片段。采用DNAMAN软件拼接, 获得全长cDNA序列(GenBank登录号为KC890769), 总长度为2015 bp, 其中5′-UTR为99 bp, 3′-UTR为182 bp, ORF为1734 bp, 编码577个氨基酸。
2.2 TaNRX基因编码蛋白的生物信息学分析 2.2.1 蛋白的二级结构及理化性质 预测该蛋白主要由α-螺旋、β-折叠、无规则卷曲、延伸链构成(表4); 理论分子量为63.79 kD, 理论等电点为4.82, 分子式为C2866H4427N729O876S21, 脂肪系数为81.47, 平均亲水性为-0.260, 可能为脂溶性蛋白, 不稳定系数为31.32, 表明该蛋白为稳定蛋白[ 21]。
表4
Table 4
表4(Table 4)
| 表4 TaNRX蛋白的二级结构预测 Table 4 Secondary structure prediction of TaNRX protein |
2.2.2 蛋白的三级结及保守结构域 预测结果表明在第39~134位、第201~294位和第364~457位有3个与锥虫氧还蛋白(tryparedoxin, TryX)类似的Trx-like结构域, 且N-端的Trx-like结构域中存在WCPPC结构, 说明该蛋白属于Trx超家族中核氧还蛋白类, 将其基因暂定名为小麦核氧还蛋白基因, 以 TaNRX表示。同时, TaNRX的C-端Trx-like结构域活性中心处氨基酸序列为WCGPC, 属于Cys- X1-X2-Cys结构, 推测其具备催化氧化还原反应的活性; 在第518~545位存在富含半胱氨酸的C1-like锌指结构, 与玉米NRX的结构一致[ 13]。
2.2.3 信号肽与跨膜结构 预测结果显示, 该蛋白的N端至第70位氨基酸之间剪切位点分值( C-value)和综合剪切分值( Y-value)无明显峰值, 说明该蛋白的N端不存在剪切位点, 表明其不包含信号肽, 推测为非分泌蛋白。显示其不具有跨膜结构, 不是膜定位的蛋白。
2.2.4 同源分析 BALSTP比对结果显示, TaNRX及其同源相似序列在单、双子叶植物间差距较大, 分别形成2个进化枝; 而在单子叶植物中, 小麦与大麦的亲缘关系最近, 同源性达93%, 与水稻的同源性仅为76% (图1)。
图1
Fig. 1
| Figure OptionViewDownloadNew Window | |
 | 图1 TaNRX与其他物种同源序列的进化树Fig. 1 Phylogenic tree of TaNRX and other homologous sequences from different species |
2.3 TaNRX基因结构和在晋麦47与西农2208之间序列差异分析 以中国春双体、缺体-四体和双端体系对引物Nra1、Nra2和Nra3进行染色体定位, 除缺体-四体N5B-T5A和双端体DT-5BL的基因组DNA的PCR没有扩增出预期特异条带外, 3对引物均在中国春的双体和其余缺体-四体和双端体扩增出预期的特异条带(图2)。证明这3对引物均位于5BS染色体上, 说明目标基因也位于5BS上。
图2
Fig. 2
| Figure OptionViewDownloadNew Window | |
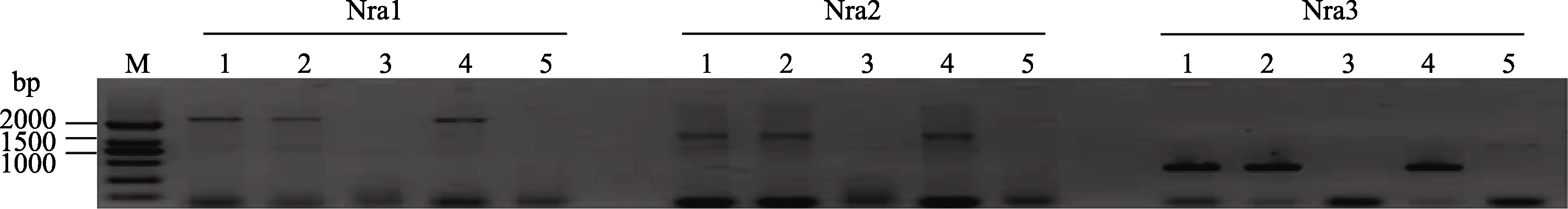 | 图2 扩增 TaNRX基因的3对引物的染色体定位M: DNA marker 2000; 1: 中国春; 2: N5A-T5B; 3: N5B-T5D; 4: N5D-T5B; 5: DT-5BL。Fig. 2 Chromosomal localization of three pairs of primers for amplification of TaNRX geneM: DNA marker 2000; 1: Chinese Spring; 2: N5A-T5B; 3: N5B-T5D; 4: N5D-T5B; 5: DT-5BL. |
以晋麦47基因组DNA为模板, 利用上述3对特异性引物进行PCR扩增和测序, 最终克隆获得了 TaNRX基因组的全长序列, 由4148 bp碱基对组成; 与cDNA序列比对发现, 该基因包含大小为90、487、489和668 bp的4个外显子和大小为1305、94和783 bp的3个内含子(图3)。在西农2208基因组中进行 TaNRX全序列克隆和测序, 并与在晋麦47基因组的克隆序列进行比较, 发现 TaNRX基因全序列在两品种间的一致性达97.0%, 存在63个单核苷酸变异(SNP)位点、6个插入(Ins)位点和8个缺失(Del)位点; 其中在第1个内含子区内存在较大差异, 包含43个SNP、4个Ins和6个Del (图3)。
图3
Fig. 3
| Figure OptionViewDownloadNew Window | |
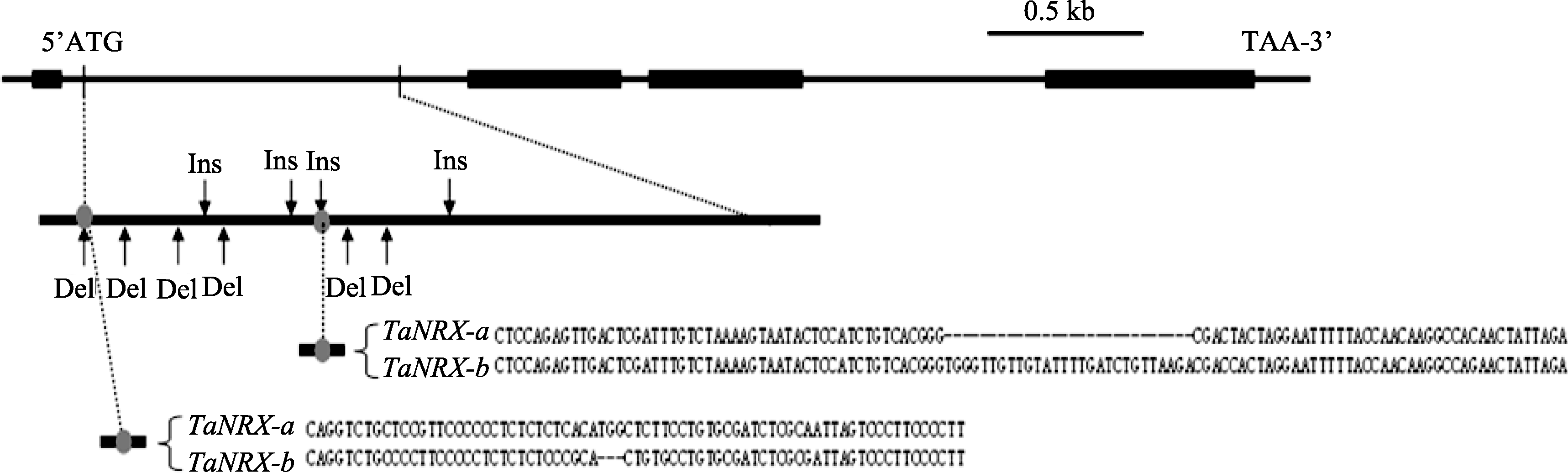 | 图3 晋麦47 ( TaNRX-a)和西农2208 ( TaNRX-b)中 TaNRX基因结构和序列变异示意图外显子用粗线表示; 非翻译区用细线表示, 包括内含子、5′-UTR和3′-UTR。Fig. 3 Sketch map of structure and sequence variation of gene TaNRXin Jinmai 47 ( TaNRX-a) and Xinong 2208 ( TaNRX-b)The thick line shows the position of exon, and the thin line shows the position of intron, 5′-UTR, or 3′-UTR. |
2.4 TaNRX与抗旱相关分子标记的开发与验证 利用相对发芽率差异明显的2组品种进行 TaNRX基因组全序列测序比较, 发现在第1内含子区段内, 抗旱性极强的鲁麦21和临旱917与晋麦47序列一致, 而抗旱性极弱的济麦20和陕150与西农2208序列一致。进而在第1内含子区1个3 bp的Del位点(图3)和紧邻的3个SNP位点, 设计两个标记 TaNRX-a1和 TaNRX-b1 (表3); 在第1内含子区1个28 bp的Ins位点(图3)设计另2个标记 TaNRX-a2和 TaNRX-b2 (表3)。4个标记引物的染色体定位结果与克隆基因所用引物一样, 均定位于5BS之上。
4个标记用于全国150份品种(系)检测, 标记 TaNRX-a1和 TaNRX-a2在鲁麦21等92份品种(系)中分别扩增出841 bp和845 bp的预期条带, 而在济麦20等58份品种(系)中均没有扩增出预期条带, 说明 TaNRX-a1与 TaNRX-a2为显性标记。标记 TaNRX-b1和 TaNRX-b2在济麦20等58份品种(系)中分别扩增出870 bp和910 bp的预期条带, 而在鲁麦21等92份品种(系)中没有扩增出任何条带, 说明 TaNRX-b1与 TaNRX-b2也为显性标记, 同时 TaNRX-a1和 TaNRX-a2分别与 TaNRX-b1和 TaNRX-b2为互补标记。依据上述结果, 将该基因分为2种等位变异类型 TaNRX-a和 TaNRX-b。
在150份品种(系)中, TaNRX-a和 TaNRX-b基因型分别为92份和58份(表5), 依次占61.3%和38.7%。 TaNRX-a基因型品种(系)平均相对发芽率(73.08%)高于 TaNRX-b基因型(44.49%), 差异极显著( P<0.01)。其中, 在相对发芽率≥90%的抗旱性极强品种(系)中, 分子标记检测均为 TaNRX-a基因型品种(系); 在相对发芽率在70.0%~89.9%之间的品种(系)中, 标记检测为 TaNRX-a的品种(系)占91%以上; 在相对发芽率≤29.9%的抗旱性极弱品种(系)中, 分子标记检测均为 TaNRX-b基因型品种(系); 在相对发芽率在30.0%~49.9%之间的品种(系)中, 检测为 TaNRX-b基因型的品种(系)占70%以上。这一结果表明分子标记检测为 TaNRX-a基因型小麦的总体抗旱性高于 TaNRX-b基因型, 4个标记可用于小麦种质资源抗旱性的鉴定筛选。
图4
Fig. 4
| Figure OptionViewDownloadNew Window | |
 | 图4 以4个分子标记检测小麦品种 TaNRX基因等位变异的电泳图谱M: DNA marker 2000; 1: 济麦20; 2: 鲁麦21; 3: 陕150; 4: 临旱917; 5: 西农2208; 6: 晋麦47; 7: 秦麦9号; 8: 洛麦8918; 9: 郑麦9023; 10: 西农8727; 11: 武农148; 12: 铜麦4号; 13: 晋麦54; 14: 天94-3; 15: 新麦26; 16: 中麦9号; 17: 泰山21。Fig. 4 Profile of electrophoresis on TaNRX allelic variants in wheat varieties using four markersM: DNA marker 2000; 1: Jimai 20; 2: Lumai 21; 3: Shaan 150; 4: Linhan 917; 5: Xinong 2208; 6: Jinmai 47; 7: Qinmai 9; 8: Luomai 8918; 9: Zhengmai 9023; 10: Xinong 8727; 11: Wunong 148; 12: Tongmai 4; 13: Jinmai 54; 14: Tian 94-3; 15: Xinmai 26; 16: Zhongmai 9; 17: Taishan 21. |
表5
Table 5
表5(Table 5)
| 表5 TaNRX-a和 TaNRX-b基因型的相对发芽率(RGR)及抗旱性不同级别的品种数 Table 5 Relative germination rate (RGR) and number of cultivars with different grades of drought resistance in TaNRX-a and TaNRX-b genotypes | |||||||||||||||||||||||||||||||||||||||||||
3 讨论3.1 小麦 TaNRX及其编码蛋白的结构特点分析 核氧还蛋白(NRX)最早由Kurooka等[ 22]发现并命名, 因其包含Trx-like结构域Cys-X1-X2-Cys活性区而被认定为Trx超家族的新成员, 并将其定位于细胞核, 植物的NRX最早在玉米中被发现[ 13]。所有的NRX蛋白在其Trx-like结构域中至少有1个WCPPC结构。相对于Trx典型的结构域而言, TaNRX中的Trx-like结构与锥虫氧还蛋白 (tryparedoxin, TryX)结构更为接近[ 23]。本研究中得到的小麦目标基因编码的蛋白序列具备NRX结构特征, 以 TaNRX表示。NCBI数据库中与小麦 TaNRX编码蛋白有较高同源性的二穗短柄草
(XP_003557743)、水稻(NP_001050329)、高粱(XP_002467709)等序列其相应的基因结构均为4个外显子和3个内含子, 与 TaNRX基因结构一致, 它们分别位于二穗短柄草的第1染色体、水稻的第3染色体、高粱的第1染色体上。前人对比较基因组学的研究指出小麦的第5染色体与二穗短柄草的第1、水稻的第3染色体等存在良好的共线性关系[ 24, 25], 本文克隆的 TaNRX基因恰好定位在小麦的第5染色体上, 说明本文分离得到的基因确为小麦 TaNRX基因。
小麦 TaNRX编码蛋白的第2个Trx-like结构域的中心序列为GYPPV, 与玉米中发现的NRX (ZmNRX)第2个Trx-like结构域的中心序列GFPPL类似, 两者均无Cys-X1-X2-Cys结构而无法形成可逆的二硫键氧化还原中心[ 13]。TaNRX的C-端的半胱氨酸能形成锌指结构, 与ZmNRX结构一致, 而明显区别于哺乳动物类的NRX, 这可能与动植物的生理代谢方式及所涉及的功能差异有关[ 13, 23]。小麦TaNRX的C-端Trx-like结构域中心序列为WCGPC, 不同于ZmNRX在其N-端与C-端均为WCPPC, 拟南芥NRX相似蛋白(GenBank登录号AAM64954)的N-端Trx-like结构域中心序列为WCGPC, C-端Trx-like结构域中心序列为WCPPC结构, 与TaNRX结构恰好相反。上述差异可能是具种属间差异的不同物种体内的不同代谢环境所引起的。
3.2 小麦 TaNRX的等位变异与抗旱性关系 基于小麦 TaNRX第1内含子区两处差异位点设计的4对引物, 检测结果相同或互补, 可以相互验证, 进一步说明设计标记所涉及的区段序列在变异上具有同步性。真核mRNA内含子能在转录与翻译等多个过程调控基因表达, 推测 TaNRX在第1内含子区的等位变异可能与调控该基因的表达有关。He等[ 26]基于黄色素合成相关的 PSY基因在内含子区的差异开发了功能分子标记, Fu等[ 27]研究春化基因 VRN-1认为 Vrn-B1、 Vrn-D1在第1个内含子区的1个大片段缺失包含了介导开花抑制的识别位点, 并开发了识别不同等位变异类型的功能分子标记。本文基于两组极端抗旱品种第1内含子区之间的差异序列开发了分子标记, 用150份抗旱性存在明显差异的品种(系)验证表明, 标记所划分的不同等位变异基因型材料的相对发芽率存在显著差异, 所开发的标记能在一定程度上区分材料的萌发期抗旱性强弱。本文标记检测为 TaNRX-a基因型品种 (系)萌发期的总体平均相对发芽率明显高于 TaNRX-b基因型, 可能与其在干旱胁迫下大量表达, 参与逆境下的胁迫信号传递和活性氧清除等功能有关。但在所检测的品种(系)中, 有6%的 TaNRX-a基因型材料萌发抗旱性较弱, 2%的 TaNRX-b基因型材料萌发抗旱性较强, 一方面干旱胁迫下种子萌发可能还受其它生化物质的调控, 另一方面可能与小麦适应干旱胁迫途径的多样性有关, 也许 TaNRX基因还存在与抗旱相关的其他等位变异类型。
目前对植物NRX功能的研究报道较少, 对其参与的代谢途径也不甚清楚。本研究基于基因型间萌发期抗旱性差异显著性分析, 表明 TaNRX基因很可能参与了小麦调控干旱胁迫的生理适应过程, 但涉及的调控机理有待进一步试验研究。干旱胁迫应答包含多个生理生化过程, 小麦在生长发育的不同阶段对干旱的耐受性差异使其抗旱性的研究更为复杂, 随着对小麦抗旱机理研究的深入, 在明确不同生育期抗旱相关主效基因的基础上开发多个功能标记, 能弥补单个基因标记的不足, 更全面地指导抗旱品种的选育。
4 结论从普通小麦中克隆了一个位于5BS染色体的Trx超家族新基因 TaNRX, 其开放阅读框由1734 bp组成, 编码577个氨基酸。小麦 TaNRX基因与抗旱相关的等位变异包括 TaNRX-a和 TaNRX-b基因型。基于萌发期相对发芽率极端品种第1内含子区域的差异序列, 开发了4个显性互补的分子标记, 用其检测表明 TaNRX-a基因型品种萌发期的平均相对发芽率明显高于 TaNRX-b基因型, 说明开发的标记可以用于小麦抗旱性筛选鉴定。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] | |
| [25] | |
| [26] | |
| [27] |

{kind=link}
{kind=link}
{kind=link}
{kind=link}