* 通讯作者(Corresponding authors): 薛吉全, E-mail:xjq2934@163.com; 郭东伟, E-mail:gdwei1973@126.com 收稿日期:2013-10-20 基金:
摘要
关键词:玉米; 胚乳细胞; 原生质体; 内多倍化; 分离; 流式分选
Isolation and Flow Purification of Endosperm Protoplast from Developing Seed of Maize
GUO Yan-Ping
Fund:
Abstract
Keyword:Maize; Endosperm; Protoplast; Endoreduplication; Isolation; Flow sorting
Show Figures
Show Figures


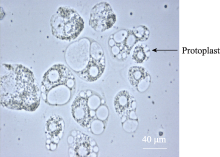

玉米是重要的工业原料作物, 对国民经济发展具有重要意义。玉米籽粒干物质的70%~90%都存储于胚乳, 是玉米经济和营养价值的主要来源, 因此胚乳的正常发育是决定玉米经济产量形成的关键因素[ 1]。和其他禾谷类作物一样, 玉米胚乳也属于核型胚乳, 是植物双受精的产物。研究表明, 在受精后2~4 h, 由源自母本的两个中央极核和一个源自父本的精核融合而成的三倍体细胞核即开始快速同步分裂, 形成由同一细胞膜包被的双核体。随后, 双核体继续分裂8~10次, 发育成一个含有256~512个细胞核的多核体。之后, 胚乳细胞内部各核之间的微管不断加厚形成细胞壁, 逐渐将每个细胞核分隔开来, 并最终发育为正常的三倍体胚乳细胞[ 2]。在玉米胚乳中, 细胞化过程通常在授粉后4 d就已经完成, 随后胚乳细胞进行正常的有丝分裂, 完成胚乳发育所需细胞数目的增长, 直至授粉后8~10 d, 胚乳中部的一些细胞开始停止有丝分裂, 继而转向一种称之为内复制(endoreduplication)的特殊的细胞分裂, 并逐渐向胚乳外部扩散。在内复制过程中, 胚乳细胞每个周期只进行一次正常的DNA复制, 但不分裂, 产生DNA加倍一至数轮的大量内复制细胞。从发育进程来看, 这种内复制分裂始于授粉后8~10 d, 16 d左右达到最大程度, 18~20 d后, 随着籽粒淀粉的积累、细胞的失水凋亡而逐渐停止。此时, 胚乳中可检测到DNA含量达96 C的内复制细胞。内复制的直接结果就是细胞体积的快速增加, 由于胚乳细胞的内复制进程与籽粒的快速膨大完全同步, 因此被认为是玉米籽粒发育和经济产量形成的关键性动力[ 3]。理解玉米胚乳细胞内复制的分子调控机理和生物学意义, 可为改善栽培措施、培育更具经济价值的玉米新品种奠定理论基础。此外, 玉米胚乳还是研究细胞发育的理想模型。根据各部分细胞的功能和特征, 可将玉米胚乳分为糊粉层(aleurone)、淀粉区(starchy endosperm, SE)、基部转化层(basal endosperm transferlayer, BETL)和胚周区(endosperm surrounding region, ESR)四部分, 尽管这些组织都由同一胚乳细胞发育而来, 却经历了不同的分化途径。玉米胚乳各部分以及整个胚乳组织细胞发育的分子调控机制至今仍不清楚。
转录组分析是新近发展起来的以DNA二代测序为基础的基因表达分析技术, 它能够在整个转录组水平上对基因的差异表达、SNP检测和可变剪切等转录水平上的表达情况快速系统分析, 有利于用系统生物学的方法描绘基因表达和调控的时空网络。目前, 该方法已广泛用于多个物种基因的表达分析[ 4], 但这些研究多集中在组织水平上, 以特定细胞为对象的转录组分析研究仍鲜有报道。如果通过特定分离技术将不同特征的胚乳细胞分离出来进行转录组分析, 就有可能对内多倍化、细胞凋亡、细胞分化等生命现象的分子调控更精确地解释。为此, 本研究在借鉴前人研究的基础上, 通过调整酶、渗透调节物质和膜稳定剂的种类、浓度及反应时间, 优化了一套玉米胚乳细胞原生质体的酶解方案, 并运用流式分选技术纯化原生质体, 以期为更深入的特定细胞转录组分析及胚乳细胞原生质体培养提供技术支撑。
1 材料与方法1.1 试验材料玉米自交系郑58, 由西北农林科技大学农业部西北旱区玉米生物学与遗传育种重点实验室提供。分期播种材料, 人工套袋授粉, 取授粉后8~ 10 d新鲜果穗于4℃保存备用。
1.2 实验方法1.2.1 原生质体制备 在超净工作台上, 用手术刀(11号)从新鲜果穗上切取若干完整籽粒, 横切, 取中部淀粉区组织于5.5 cm一次性培养皿, 切成1 mm左右小块, 转入2.0 mL离心管, 确保总量控制在0.4~0.6 g之间, 加酶解液1 mL, 振荡混匀, 使酶液与组织充分接触, 于30℃, 40转min-1振荡培养2~ 14 h, 每隔2 h取样1次, 统计原生质体数, 确定最佳酶解时间。待酶解完成后, 以巴斯德吸管吸取酶解混合液, 100 μm滤网过滤, 以血球计数板在显微镜下计算原生质体产量和活力。酶解液以MS为基础培养基, pH 5.8, 采用3组合酶, 每种酶设高低2个浓度, 分别为纤维素酶0.5%、1.0%; 半纤维素酶0.5%、1.0%; 离析酶0.3%、0.5%, 共8个处理(表1)。设0.6、0.7、0.8、0.9 mol L-1 4个渗透压稳定剂蔗糖浓度处理; 质膜稳定剂处理分别为Z1: 80% CaCl2+20% MES+0.5 mmol L-1 DTT; Z2: 100% CaCl2+25% MES+0.5 mmol L-1 DTT; Z3: 120% CaCl2+30% MES+0.5 mmol L-1 DTT; Z4: 150% CaCl2 +35% MES +0.5 mmol L-1 DTT。
1.2.2 流式纯化 酶解后的原生质体混合物, 加等量无酶培养液稀释1倍, 经100 μm滤网过滤, 滤液加二乙酸荧光素(FDA)染液, 终浓度0.01%, 避光常温染色30 min。用流式细胞仪(FACS calibur, Becton Dickinson, USA)分选, 分选参数为输出功率150 mW, 电压400 V, 激发波长488 nm, 指数放大,
表1
Table 1
表1(Table 1)
| 表1 不同酶浓度处理组合 Table 1 Treatment combination with different enzyme concentrations (%) |
分析流速1000~2000 Particle s-1。散点图设门分选, 分选模式Normal-R, 分选流速100~150 Particle s-1。以CEllquest软件获取数据进行流式核型分析。将原生质体直接分选至干净载玻片上, 以荧光倒置显微镜(Nikon, TE 2000-5)检测分选原生质体活力。
1.2.3 原生质体产量与活力统计 (1)原生质体产量测定(采用血球计数板计数), 吸取原生质体混合滤液10 μL, 悬浮于血球计数板上, 在显微镜下逐次数出16个方格内的原生质体。每样品重复3次, 取平均值; (2)原生质体活力测定(FDA染色法), 用丙酮配成5 mg mL-1 FDA溶液, 按照25 μL mL-1 FDA对原生质体染色, 染色后放于黑暗处20 min。在荧光显微镜下, 有活力的原生质发出绿色荧光, 无活力的不发光。随机选取5个视野进行统计, 每样品重复3次。原生质体存活率=(有活力的原生质体数/原生质体总数)×100%。
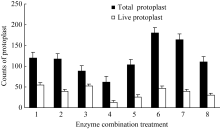
2 结果与分析2.1 原生质体分离2.1.1 不同酶浓度及组合对原生质体产量和活力的影响 原生质体以血球记数板随机统计12个记数区域内的数目显示, 不同酶处理对原生质体的产量和活力有显著影响。以8种酶组合分别处理胚乳组织过夜酶解后, 结果如图1所示, 处理6原生质体产量最高, 达到180个, 其中有活力原生质体47个, 占总数的26.1%。处理4产量最低, 仅62个, 其中活原生质体13个, 占21.0%。不同酶浓度处理对原生质体产量及活力的影响也存在明显差异。在3种酶中, 离析酶对原生质体产量影响最大, 0.5%浓度下的原生质体总数比0.3%水平增加45%, 而原生质体活力没有显著下降。纤维素酶则对原生质体活力影响最大, 低浓度水平(0.5%)下有活力原生质体占原生质体总数的百分比为36.3%, 显著高于高浓度水平(1.0%)下的27.5%; 而在两种浓度纤维素酶水平下, 原生质体总数没有显著差异。半纤维素酶对原生质体总数和原生质体活力的影响介于纤维素酶和离析酶之间, 低浓度水平更有助于原生质体总数的提高和原生质体活性的维持, 半纤维素酶的这种影响同时受纤维素酶和离析酶的影响, 在离析酶高浓度水平下, 配合高浓度的纤维素酶比低浓度更有助于提高原生质体产量和活力。综上认为, 酶组合处理6 (0.5%的离析酶+0.5%半纤维素酶+1%纤维素酶)为玉米胚乳原生质体分离的最佳选择。
图1
Fig. 1
| Figure OptionViewDownloadNew Window | |
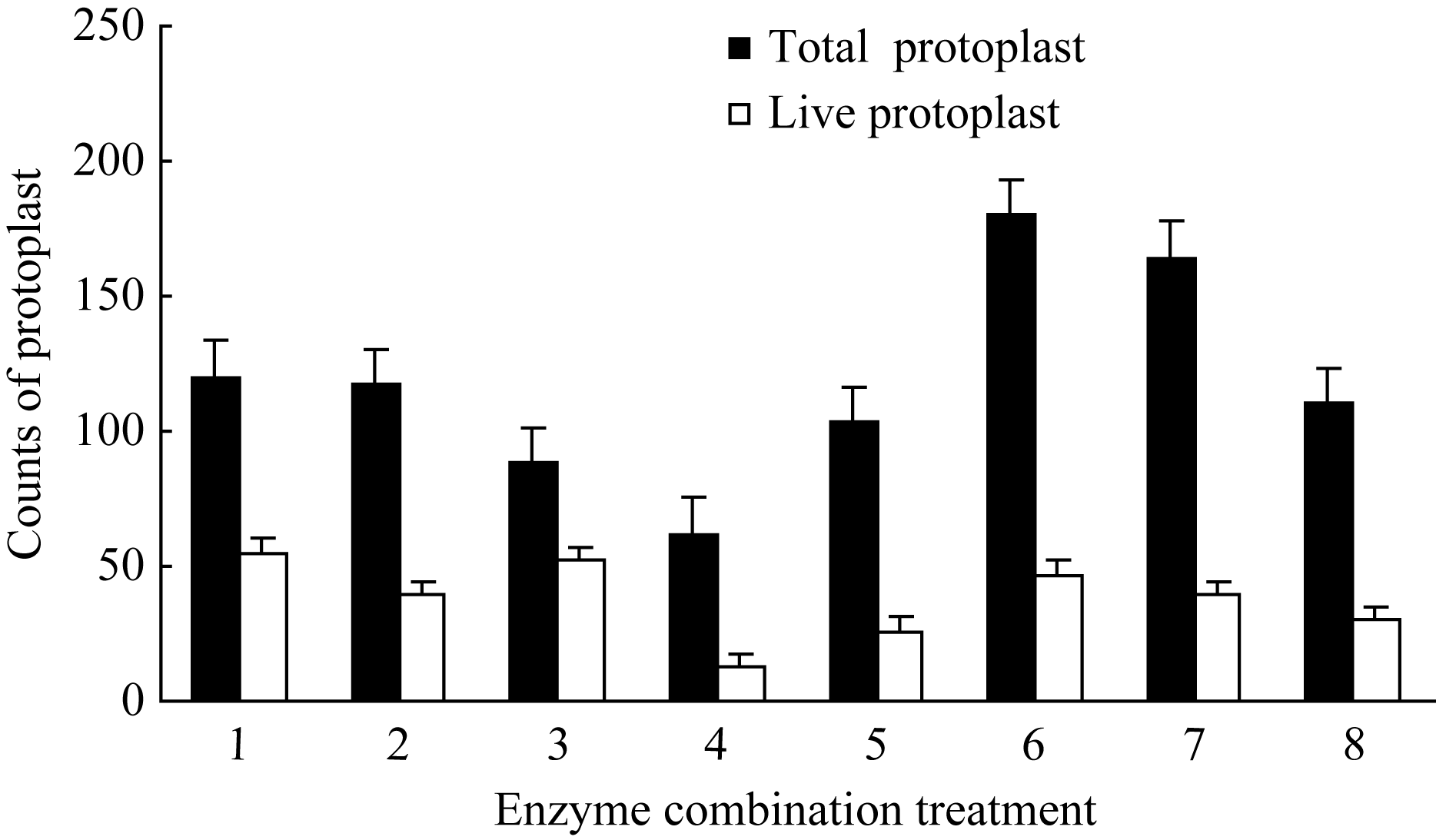 | 图1 不同酶浓度及组合对原生质体产量与活力的影响1: 0.3%离析酶+0.5%纤维素酶+0.5%半纤维素酶; 2: 0.3%离析酶+1.0%纤维素酶+0.5%半纤维素酶; 3: 0.3%离析酶+0.5%纤维素酶+1.0%半纤维素酶; 4: 0.3%离析酶+1.0%纤维素酶+1.0%半纤维素酶; 5: 0.5%离析酶+0.5%纤维素酶+0.5%半纤维素酶; 6: 0.5%离析酶+1.0%纤维素酶+0.5%半纤维素酶; 7: 0.5%离析酶+0.5%纤维素酶+1.0%半纤维素酶; 8: 0.5%离析酶+1.0%纤维素酶+1.0%半纤维素酶。Fig. 1 Effect of different enzyme concentrations and composition on the yield and vigor of protoplasts1: 0.3% macerozyme+0.5% cellulase+0.5% hemicellulase; 2: 0.3% macerozyme+1.0% cellulase+0.5% hemicellulase; 3: 0.3% macerozyme+0.5% cellulase+1.0% hemicellulase; 4: 0.3% macerozyme+1.0% cellulase+1.0% hemicellulase; 5: 0.5% macerozyme+0.5% cellulase+0.5% hemicellulase; 6: 0.5% macerozyme+1.0% cellulase+0.5% hemicellulase; 7: 0.5% macerozyme+0.5% cellulase+1.0% hemicellulase; 8: 0.5% macerozyme+1.0% cellulase+1.0% hemicellulase. |
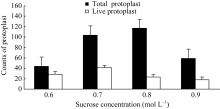
2.1.2 渗透压稳定剂对原生质体的影响 维持适当的渗透压是确保细胞正常代谢的基本条件之一。细胞内外的渗透压差决定着细胞与外部环境之间的物质交流取向。当渗透压差过大时细胞将丧失主动吸收能力, 造成水分缺失或过渡吸水, 给细胞的空间结构带来严重伤害, 进而影响到细胞活力。在原生质体分离过程中, 由于失去了细胞壁的保护, 原生质体会更加脆弱, 因此确保反应体系与原生质体内渗透压平衡对于原生质体分离的产量和活力至关重要[ 5]。本文以蔗糖为渗透压调节剂, 研究了4种浓度下, 以最佳酶处理组合处理胚乳组织过夜酶解时, 原生质体时产量和活力。图2表明, 不同渗透压水平下, 原生质体产量和活力存在明显差异; 原生质体产量和活力随渗透压的增大而提高, 但二者的峰值并不同步, 在0.8 mol L-1时, 16个计数区域内原生质体总数117个, 达到最高, 而有活力的原生质体只有23个; 原生质体活力则在0.7 mol L-1维持了最高水平, 在统计的104个原生质体中, 有活力原生质体41个, 占总数的39.4%; 因此, 在玉米胚乳的原生质体分离时, 酶解液蔗糖浓度应介于0.7~0.8 mol L-1之间, 更高浓度(0.9 mol L-1)蔗糖处理下, 导致原生质体产量和活力显著下降。这可能是因为低浓度 (0.6 mol L-1)水平时, 原生质体渗透压远大于酶分离液的渗透压, 细胞被动吸水胀破或结构发生改变。在高蔗糖浓度水平下, 原生质体渗透压低于分离液, 细胞失水皱缩, 部分原生质体因体积变小在过滤时流失, 另一部分原生质体则因为失水失活, 最终导致原生质体产量和活力下降。
图2
Fig. 2
| Figure OptionViewDownloadNew Window | |
 | 图2 渗透压稳定剂对原生质体分离的影响Fig. 2 Effect of osmotic pressure regulator on protoplasts isolation |
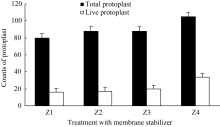
2.1.3 质膜稳定剂对原生质体的影响 与渗透压稳定剂作用类似, 在酶解反应体系中加入质膜稳定剂也是为了保护原生质体膜的完整性或促进原生质体培养时细胞壁的再生[ 6], 常用的质膜稳定剂有葡萄糖硫酸钾、MES、氯化钙等[ 7]。本文以氯化钙和MES为质膜稳定剂, 研究了4种不同浓度配比下, 原生质体酶解的产量及活力。结果表明(图3), 在设定的4个浓度范围内, 伴随着质膜稳定剂浓度的提高, 原生质体的产量及活力均显著提高, 在Z4水平(150% CaCl2+35% MES +0.5 mmol L-1 DTT)达到峰值。至于更高浓度的质膜稳定剂能否进一步提高原生质体产量和活力, 尚待进一步实验研究。
图3
Fig. 3
| Figure OptionViewDownloadNew Window | |
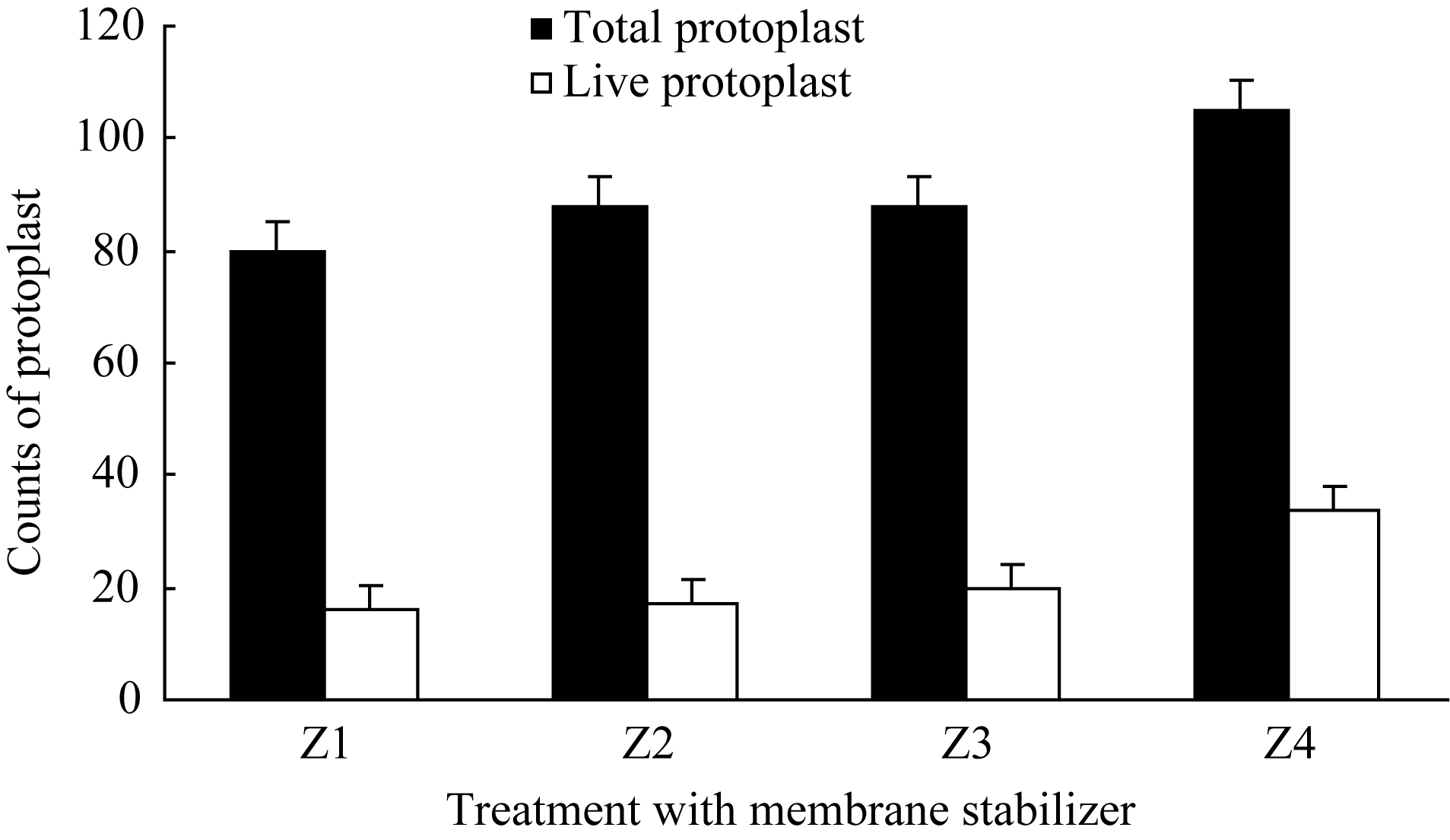 | 图3 质膜稳定剂对原生质体分离的影响Fig. 3 Effect of membrane stabilizer on protoplasts isolationZ1: 80% CaCl2+20% MES+0.5 mmol L-1 DTT; Z2: 100% CaCl2+25% MES +0.5 mmol L-1 DTT; Z3: 120% CaCl2+30% MES+0.5 mmol L-1 DTT; Z4: 150% CaCl2+35% MES +0.5 mmol L-1 DTT. |
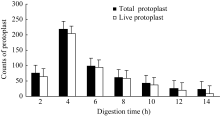
2.1.4 酶解时间对原生质体的影响 按照最佳酶处理组合、最适渗透压、最适质膜稳定剂浓度, 重新配置酶解液, 调节pH至5.8, 分装在2.0 mL离心管中, 每管1 mL, 加胚乳材料0.5 g。在30℃、40转 min-1条件下酶解, 间隔2 h取样统计原生质体总数及活原生质体数, 直至14 h。结果如图4所示, 原生质体产量和活力与酶解时间密切相关, 以酶解液处理2 h, 时间不足, 材料不能得到充分酶解, 原生质体产量较低, 12个视野内的原生质体数仅76个, 其中有活力原生质体65个, 占原生质体总数的85.5%; 4 h处理时的原生质体产量达到最大值219个, 同期的活原生质体数204个, 占总数的93.2%;处理时间超过4 h, 原生质体总数急剧降低(6~12 h), 但此时活性原生质体数占原生质体总数的百分比仍在76.9%~95.2%之间, 这说明酶解时间过长时, 源自振荡培养的物理剪切力会导致原生质体大量破裂, 产量下降, 但未破裂的原生质体在反应体系中仍能维持较长时间活性[ 8]。对于玉米胚乳材料, 4 h为最佳酶解时间。
图4
Fig. 4
| Figure OptionViewDownloadNew Window | |
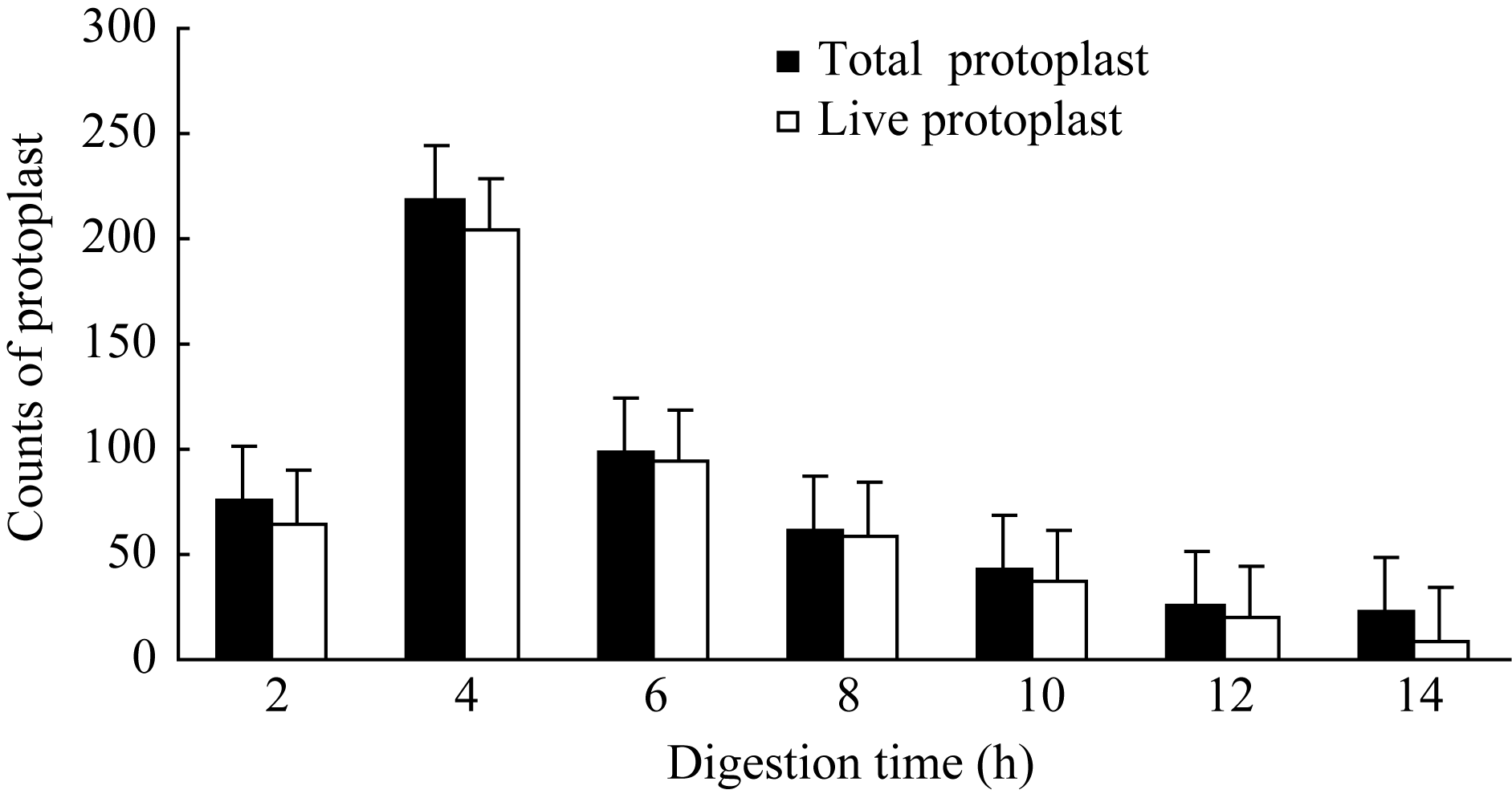 | 图4 酶解时间对原生质体分离的影响Fig. 4 Effect of digestion time on protoplasts isolation |
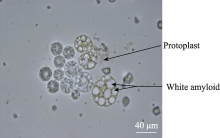
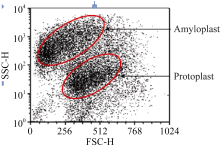
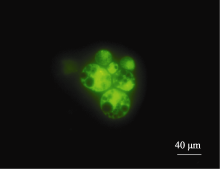
2.2 原生质体纯化酶解后的原生质体悬浮液中混有多种成分, 主要是淀粉体、原生质体和细胞碎片, 其中的淀粉体呈白色球体(图5)。淀粉体最初被细胞膜包被在细胞内, 在分离过程中由于振荡的物理剪切力或其他原因导致原生质体破裂时被释放到反应体系中, 由于一个原生质体中包含有多个淀粉体, 因此释放出的淀粉体数量常常超过体系中原生质体的数量。这些淀粉体对原生质体的后续利用会产生较大影响, 需要将它们与原生质体有效分离。我们以FDA对原生质体悬浮液染色, 采用流式细胞仪对原生质体分选纯化[ 9]。体系中有活力的原生质体能够被染色, 在荧光显微镜下被激发出绿色荧光, 没有活性的淀粉体以及失活的原生质体则不能被染色, 在显微镜下呈黑色(图6)。当染色后的原生质体悬浮液通过流式细胞仪时, 一元流式核型图上能够形成两个明显的峰(图7), 在散点图上则形成明显的2群粒子, 经分选鉴定它们分别为原生质体和淀粉体粒子群(图8), 再对原生质体群设门分选, 分离出的原生质体纯度超
图5
Fig. 5
| Figure OptionViewDownloadNew Window | |
 | 图5 未纯化原生质体Fig. 5 Unpurified protoplasts |
图6
Fig. 6
| Figure OptionViewDownloadNew Window | |
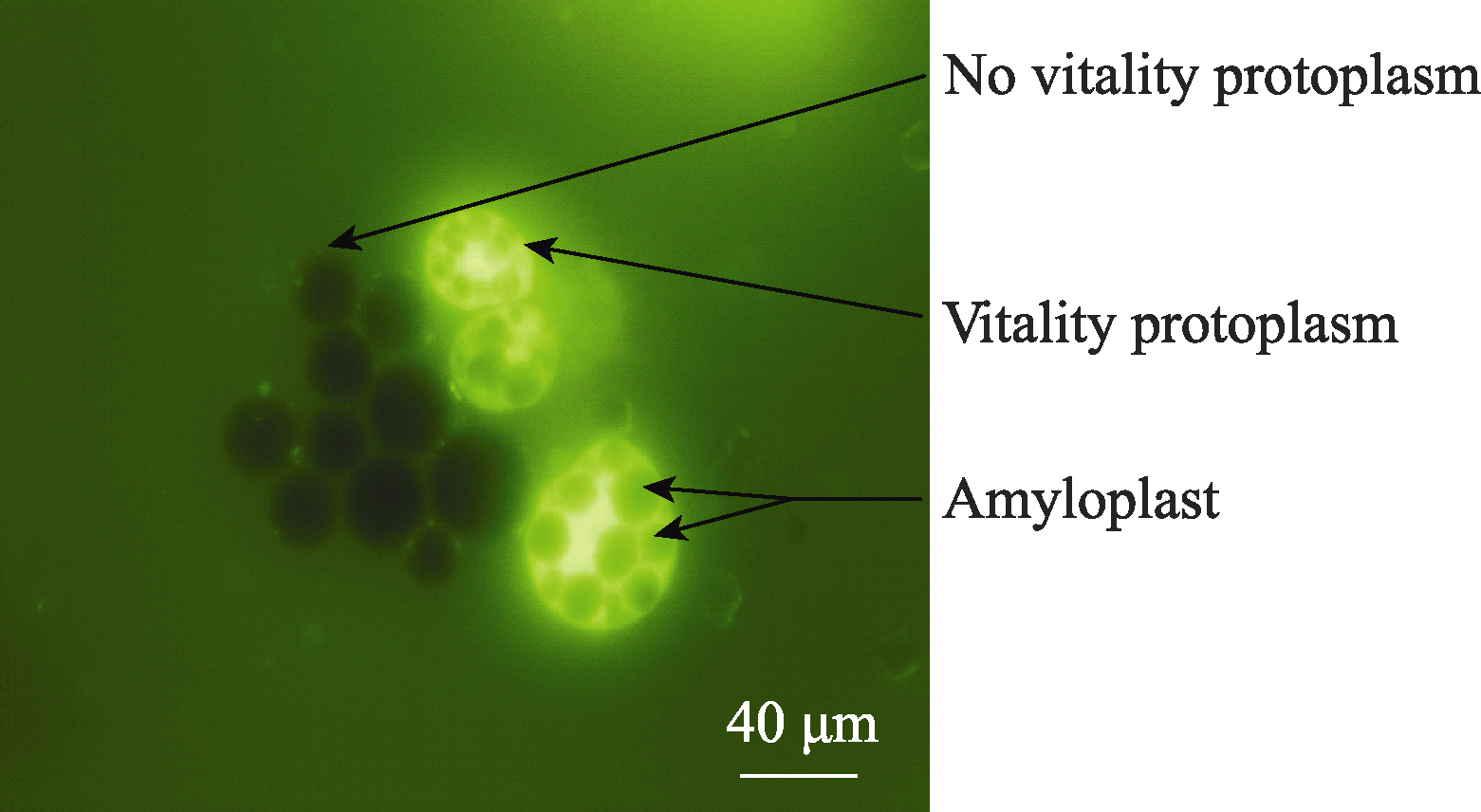 | 图6 FDA染色后的原生质体Fig. 6 Protoplasts dyed with FDA |
图7
Fig. 7
| Figure OptionViewDownloadNew Window | |
 | 图7 一元流式核型图Fig. 7 Unitary Diagram of flowing karyo |
图8
Fig. 8
| Figure OptionViewDownloadNew Window | |
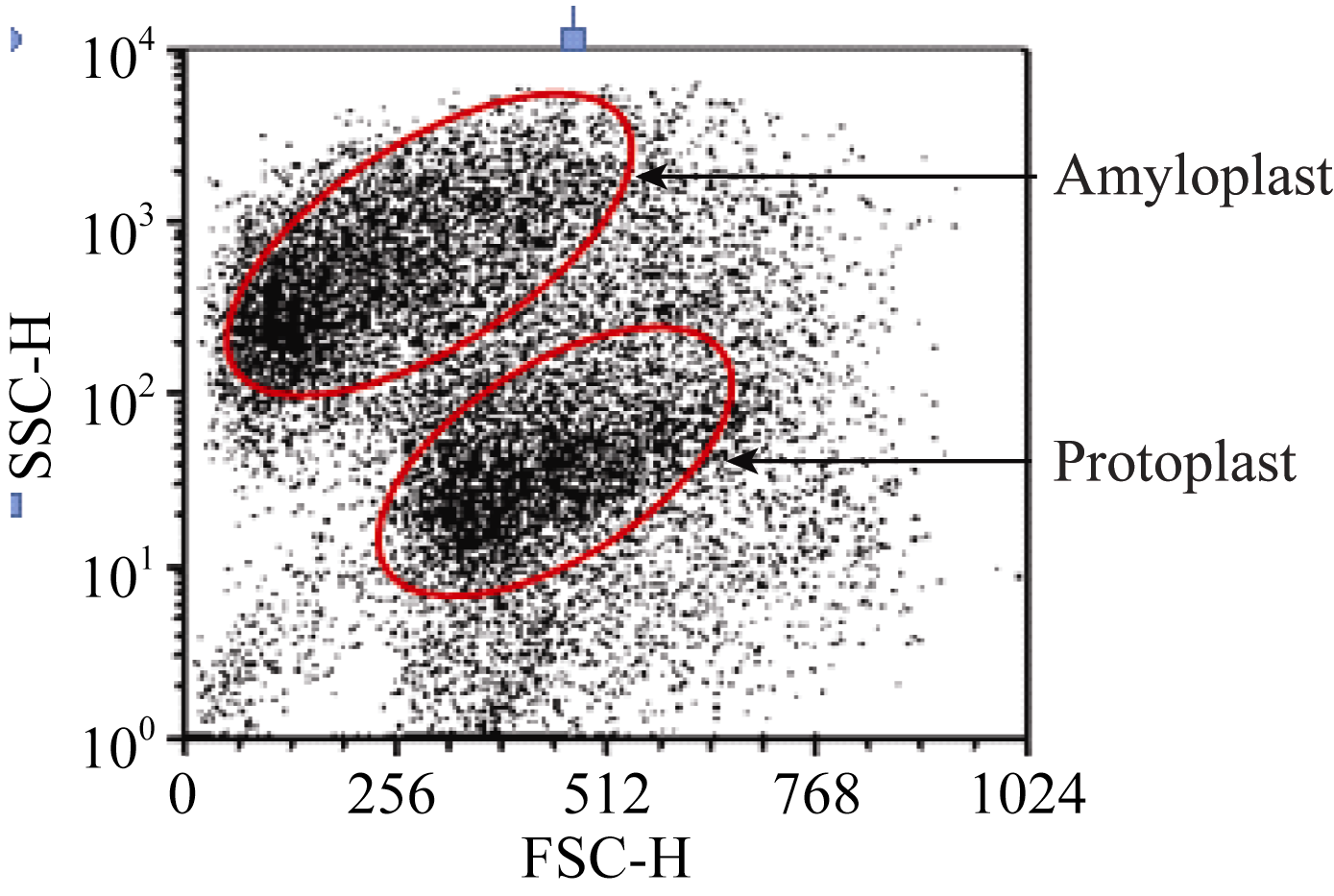 | 图8 流式散点图Fig. 8 Flowing diagram of scatter plot |
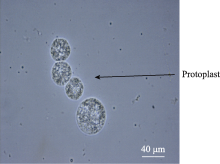
过98%, 100%保持活性(图9)。结果表明, 用流式分选法可有效分离胚乳细胞原生质体。
图9
Fig. 9
| Figure OptionViewDownloadNew Window | |
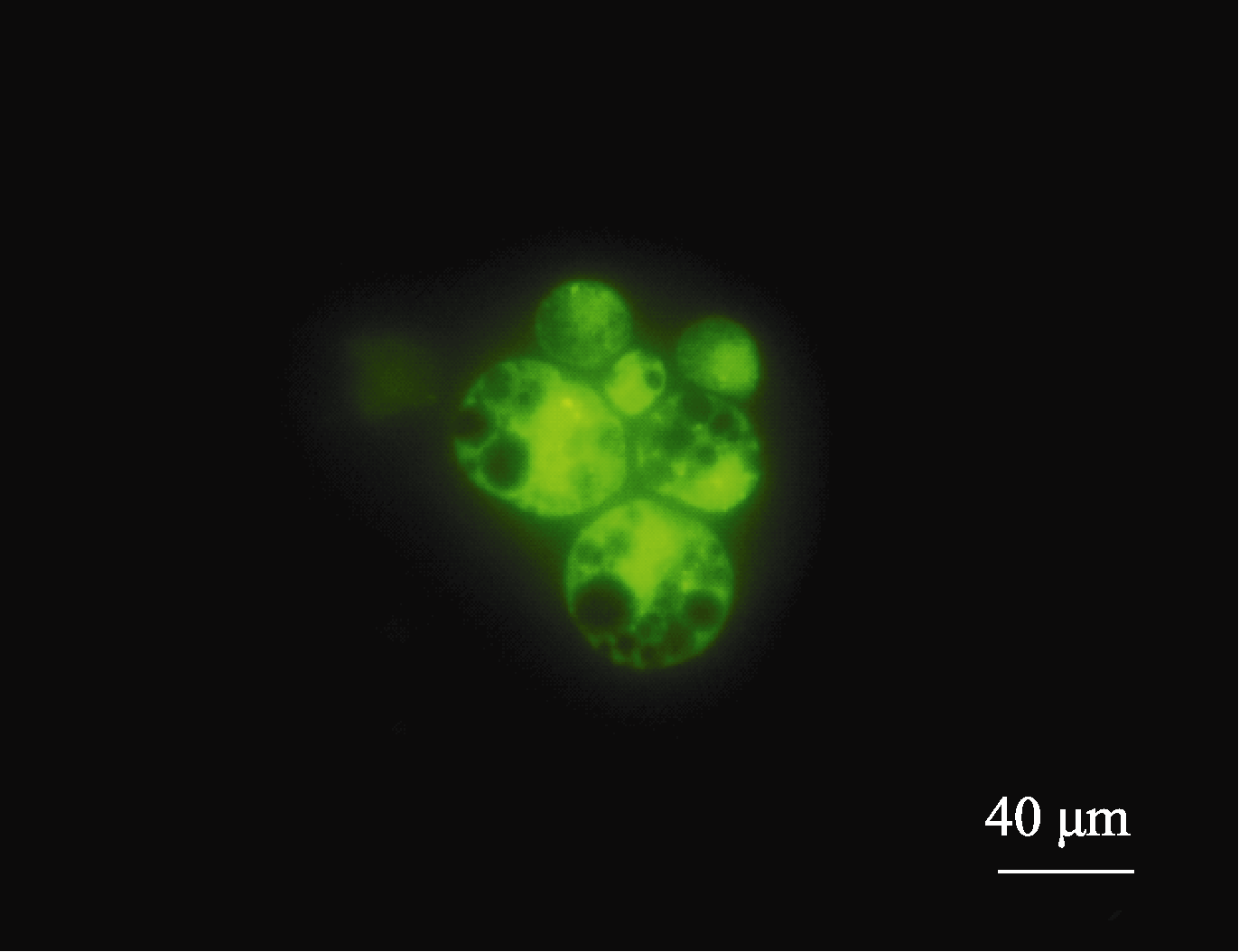 | 图9 流式纯化后的原生质体Fig. 9 Protoplasts flow sorted |
3 讨论3.1 原生质体分离原生质体是指细胞中除了细胞壁以外的所有部分, 由于没有了细胞壁的阻隔和组织结构的限制, 原生质体具有普通植物细胞所不具有的用途, 如用于体细胞杂交创造新物种及新的种质材料[ 10]; 用作转化受体进行外源遗传物质的转化以及进行细胞生长发育的理论研究[ 11]。从1960年Cooking首创了酶法分离原生质体的方法以来[ 12], 大量植物原生质体的分离、离体培养及体细胞杂交等研究获得了成功[ 13], 但这主要针对双子叶植物, 用于原生质体分离的材料也主要限制在叶肉、根尖等组织[ 14]。禾谷类作物的原生质体分离和后续利用研究进展相对较慢, 实验结果重复性较差, 仅有水稻[ 14]、大麦[ 15]等作物原生质体分离和再生的报道。玉米的原生质体分离与再生的研究一直进展缓慢, 从1987年蔡启贵等首次培养花药胚性愈伤组织原生质体并获得再生植株以来[ 16], 玉米原生质体的再生依然鲜有成功报道。理论上, 玉米各生活组织都能够通过酶解的方法获得原生质体, 但这并不包括胚乳组织, 尽管利用胚乳制备出的原生质体可为获得三倍体植株再生、胚乳细胞生长发育的分子调控、胚乳细胞表达分析等研究提供均一、稳定的受体, 但直接从幼嫩的胚乳中分离原生质体是很困难的[ 17], 这主要是因为10 d左右的胚乳细胞中积累了大量的淀粉体, 酶解处理时, 这些淀粉体在剪切力及酶解液的作用下会分散到整个反应体系, 严重限制胚乳细胞原生质体后续利用。本文参考了一些在大麦、水稻原生质体分离、玉米造粉体分离[ 18]上的方法, 将纤维素酶、半纤维素酶和果胶酶按一定浓度配比分离花后10 d左右的胚乳组织原生质体, 结果表明, 酶解处理时间对原生质体的产量及活力影响最大, 在最初过夜培养确定酶浓度及酶解反应条件时, 原生质体的产量和活力远少于在最佳处理时间4 h下的产量和活力; 用于胚乳原生质体分离的酶浓度、酶解反应条件、以及渗透压和质膜稳定剂的使用方法与前人在大麦等胚性愈伤组织酶解中所采用的方法基本一致。
3.2 原生质体纯化粗制的原生质体溶液中含有大量的淀粉体及细胞碎片(图5), 需要对原生质体分离纯化。本实验首先采用Ficoll漂浮法[ 19]去除粗酶解液中的大部分杂质(图10), 但在除杂的同时, 原生质体的数量也损失较大, 直接影响后续实验, 而采用流式纯化法, 并得到了较好的效果(图9)。
图10
Fig. 10
| Figure OptionViewDownloadNew Window | |
 | 图10 Ficoll漂浮法纯化后的原生质Fig. 10 Purified protoplasts with Ficoll flotation method |
流式细胞分选术是近年来新发展起来的一项悬浮粒子高通量分离技术, 已被广泛用于染色体、干细胞、原生质体等的分选[ 4], 但用于玉米胚乳细胞原生质体的分选尚属首次。先前的大量非特异性分选研究多采用DAPI及PI等DNA插入型荧光剂进行染色, 其中DAPI显示了较高的分辨率, 可用于植物染色体的分选。但在本研究中, 我们以DAPI对原生质体染色时, 不同原生质体间染色不均匀(图11和图12), 部分原生质体只在细胞核部位染色, 这可能是细胞膜的选择性吸收所引起的。这种细胞核染色和全细胞染色将增加分选的背景噪音, 降低细胞的分选精度, 在含有大量淀粉体背景的情况下, 以DAPI为染料可能无法对有活力原生质体有效分离。FDA染液自身无荧光, 但当它被活细胞选择性地吸入细胞后, 能被细胞质中的酯酶降解而发出荧光, 是一种有效的活细胞染色剂, 当以FDA对酶解的玉米胚乳原生质体染色时, 有活力原生质体能够被染色, 在紫外激光激发下发出绿色荧光, 失活原生质体则不能被有效染色呈黑色(图6), 染色后的原生质
图11
Fig. 11
| Figure OptionViewDownloadNew Window | |
 | 图11 未染色原生质体Fig. 11 Protoplasts un-dyed |
图12
Fig. 12
| Figure OptionViewDownloadNew Window | |
 | 图12 DAPI染色后的原生质体Fig. 12 Protoplasts dyed with DAPI |
体能被流式细胞仪识别, 将活性原生质体分离出来, 这将为后续的胚乳原生质体大规模分选以及更深入的胚乳细胞生长发育调控研究奠定基础。
4 结论授粉后8~10 d的玉米胚乳组织以1%纤维素酶, 混合0.5%的离析酶和0.5%的半纤维素酶, 在渗透压0.7~0.8 mol L-1和质膜稳定剂0.8 mol L-1的MS培养液内, 30℃消解4 h, 能够获得大量完整的粗制原生质体, 并保持90%以上的生活力。用流式分选术可将有活力原生质体从粗制原生质体悬浮液中有效富集、纯化。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}