关键词:CBRLK; SCMRP; 转基因苜蓿; 氨基酸含量; 耐碱性 Over-expressingGsCBRLK/SCMRP Enhances Alkaline Tolerance and Methionine Content in TransgenicMedicago sativa ZHAO Yang, ZHU Yan-Ming, BAI Xi, JI Wei, WU Jing, TANG Li-Li, CAI Hua* Key Laboratory of Agricultural Biological Functional Genes, Northeast Agricultural University, Harbin 150030, China Fund: AbstractDrought and saline-alkaline stresses, significantly affect growth and productivity of plants. The reaction of plant to environmental stresses is controlled by numerous genes via transcriptional regulation and protein phosphorylation. A stress-responsive kinase gene,GsCBRLK, has been cloned from aGlycine soja cDNA library under salinity, drought, and cold stresses. Over-expression ofGsCBRLK in transgenicArabidopsis resulted in enhanced tolerance to high salinity and ABA.SCMRP is a kind of storage protein gene coding high-sulfur-containing amino acid. In this study, we constructed an expression vector BEOCBRLK-SCMRP and transformed bothGsCBRLK andSCMRP genes into alfalfa. The transgenic alfalfa grew well after NaHCO3 treatment (100 and 150 mmol L-1) for 14 days; whereas, the wild type plants exhibited discoloration and stunted growth, even death. The MDA content and relative membrane permeability caused by alkaline stress in transgenic plants varied significantly compared to those in the wild type (P< 0.05). Moreover, the superoxide dismutase (SOD) activity in transgenic plants under alkali stress increased than that of the wild type. Amino acid content assay showed that the transformants had higher methionine content than the non-transformed plants. These results indicated that the transgenic alfalfa carrying bothGsCBRLKandSCMRP possesses enhanced alkaline tolerance and rich methionine simultaneously.
Keyword:CBRLK; SCMRP; Transgenic alfalfa; Methionine content; Alkaline tolerance Show Figures Show Figures
图6 NaHCO3胁迫对转基因苜蓿株高和生物量的影响柱形上不同字母表示相同处理条件下株系间有显著差异( P<0.05)。Fig. 6 Changes of plant height and biomass of transgenic alfalfa under NaHCO3 stressDifferent letters above columns indicate significant difference among lines under the same condition ( P<0.05).
图7 转基因苜蓿的MDA含量及相对质膜透性柱形上不同字母表示相同处理条件下株系间有显著差异( P<0.05)。Fig. 7 MDA content and relative membrane permeability of transgenic alfalfaDifferent letters above columns indicate significant difference among lines under the same condition ( P<0.05).
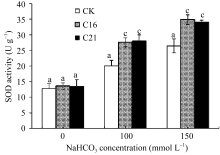
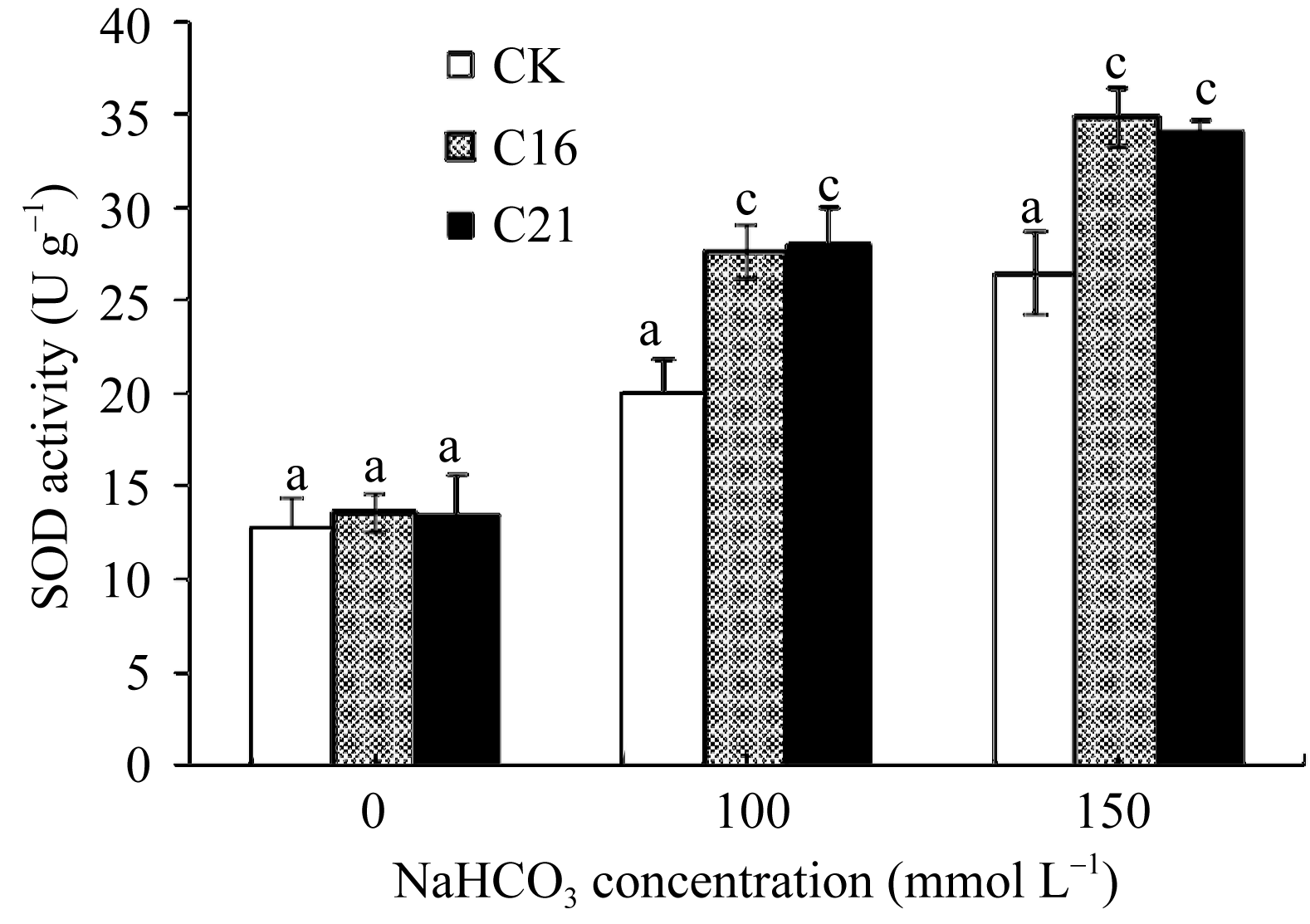
图8 碱胁迫下转基因苜蓿的SOD活性标以不同字母表示在 P<0.01水平上差异显著。Fig. 8 SOD activity in transgenic plant under NaHCO3 stressBars superscripted by different letters are significantly different at the 0.01 probability level.
表1 转基因苜蓿叶片氨基酸含量 Table 1 Content of amino acids in transgenic alfalfa (%)
氨基酸 Amino acid
NT
CS-16
CS-21
Asp
6.91±0.39
5.88±0.85*
5.89±0.54
Thr
6.48±0.00
6.08±0.21
5.97±0.10
Ser
3.65±0.05
3.52±0.08
3.54±0.11
Glu
8.74±0.09
8.70±0.08
8.58±0.16
Gly
19.28±0.20
19.14±0.55
19.79±0.74
Ala
9.40±0.07
9.44±0.12
9.58±0.25
Cys
3.88±0.05
3.83±0.08
3.91±0.11
Met
0.97±0.04
2.50±0.24**
2.10±0.68*
Ile
4.16±0.10
4.06±0.08
4.10±0.06
Leu
8.38±0.10
8.44±0.17
8.19±0.21
Tyr
1.53±0.27
1.29±0.09
1.28±0.75
Lys
6.98±0.10
7.02±0.21
7.12±0.06
His
3.49±0.09
3.62±0.05
3.52±0.05
Arg
4.96±0.03
5.08±0.14
4.82±0.14
Pro
9.51±0.01
9.40±0.36
10.00±0.32
和**分别代表转基因株系(CS-16或CS-21)与未转基因对照(NT)之间有显著( P<0.05)和极显著( P<0.01)差异。 *and** indicate significant difference between the transgenic line (CS-16 or CS-21) and the non-transformed control (NT) at P< 0.05 and P< 0.01 levels, respectively.
表1 转基因苜蓿叶片氨基酸含量 Table 1 Content of amino acids in transgenic alfalfa (%)
耿华珠. 中国苜蓿. 北京: 中国农业出版社, 1995. pp1-5GengH Z. Alfalfa in China. Beijing: China Agriculture Press, 1995. pp1-5(in Chinese)[本文引用:1]
[2]
SamacD A, SmigockiA C. Expression of oryzacy statin I and II in alfalfa increases resistance to the root-lesion nematode. Phytopathology, 2003, 93: 799-804[本文引用:1]
[3]
HipskindJ D, PaviaN L. Constitutive accumulation of resveratrol glucoside in transgenic alfalfa increases resistance to Phoma medicaginis. Mol Plant Microbe Interact, 2000, 13: 551-562[本文引用:1][JCR: 4.307]
[4]
GuoD G, ChenF, WheelerJ, WinderJ, SelmanS, PetersonM, DixonR A. Improvement of in-rumen digestibility of alfalfa forage by genetic manipulation of lignin O-methl transferases. Transgenic Res, 2001, 10: 457-464[本文引用:1][JCR: 2.609]
[5]
StrizhovN, KellerM, MathurJ, Koncz-KálmánZ, BoschD, PrudovskyE, SchellJ, SnehB, KonczC, ZilbersteinA. A synthetic cryIC gene, encoding a Bacillus thuringiensis delta- endotoxin, confers Spodoptera resistance in alfalfa and tobacco. Proc Natl Acad Sci USA, 1996, 93: 15012-15022[本文引用:1][JCR: 9.737]
[6]
ThomasJ C, WasmanmC C, EchtC, DunnR L, BohnertH J, MccoyT J. Introduction and expression of an insect proteinase inhibitor in alfalfa Medicago sativa L. Plant Cell Rep, 1994, 14: 31-36[本文引用:1][JCR: 2.509]
[7]
NarvaezV J. Orozco-Cardenas M L, Ryan C A. Differential expression of a chimeric CaMV-tomato proteinase Inhibitor I gene in leaves of transformed nightshade, tobacco and alfalfa plants. Plant Mol Biol, 1992, 20: 1149-1157[本文引用:1][JCR: 3.518]
[8]
DonnG, TischerE, SmithJ A, GoodmanH M. Herbicide resistant alfalfa cells: an example of gene amplication in plants. Mol Appl Genet, 1984, 2: 621-635[本文引用:1][JCR: 1.717]
[9]
D’HalluinK, BottermanJ, deGreef W. Engineering of herbicide-resistant alfalfa and evaluation under field conditions. Crop Sci, 1990, 30: 866-871[本文引用:1][JCR: 1.513]
[10]
PadgetteS R, KolaczK H, DelannayX, ReD B, LaValleeB J, TiniusC N, RhodesW K, OteroY I, BarryG F, EichholtzD A, PeschkeV M, NidaD L, TaylorN B, KishoreG M. Development, identification, and characterization of a glyphosate-tolerant soybean line. Crop Sci, 1995, 35: 1451-1461[本文引用:1][JCR: 1.513]
[11]
秦智慧, 晁跃辉, 杨青川, 康俊梅, 孙彦, 王凭青, 龙瑞才. 紫花苜蓿锌指蛋白基因RNAi表达载体的构建及在苜蓿的转化. 作物学报, 2010, 36: 596-601QinZ H, ChaoY H, YangQ C, KangJ M, SunY, WangP Q, LongR C. Construction and transformation of RNAi vector of MsZFN gene from alfalfa (Medicago sativa L. ). Acta Agron Sin, 2010, 36: 596-601 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[12]
安宝燕, 罗琰, 李加瑞, 乔卫华, 张宪省, 高新起. 紫花苜蓿Na+/H+逆向转运蛋白基因在拟南芥中表达提高转基因植株的耐盐性. 作物学报, 2008, 34: 557-564AnB Y, LuoY, LiJ R, QiaoW H, ZhangX S, GaoX Q. Expression of a vacuolar Na+/H+ antiporter gene of alfalfa enhances salinity tolerance in transgenic arabidopsis. Acta Agron Sin, 2008, 34: 557-564 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[13]
WinicovI. Alfin1 transcription factor overexpression enhances plant root growth under normal and saline conditions and improves salt tolerance in alfalfa. Planta, 2002, 210: 416-422[本文引用:1][JCR: 3.347]
[14]
TesfayeM, TempleS J, AllanD L, VanceC P, SamacD A. Overexpression of malate dehydrogenase in transgenic alfalfa enhances organic acid synthesis and confers tolerance to aluminum. Plant Physiol, 2001, 127: 1834-1844[本文引用:1][JCR: 6.555]
[15]
SamacD A. The influence of organic acid exudation in alfalfa on aluminum tolerance, nutrient acquisition and bacterial diversity. ISB News Rep, 2003, 11: 6-8[本文引用:1]
[16]
YangL, JiW, ZhuY M, GaoP, LiY, CaiH, BaiX, GuoD J. GsCBRLK, a calcium/calmodulin-binding receptor-like kinase, is a positive regulator of plant tolerance to salt and ABA stress. J Exp Bot, 2010, 61: 2519-2533[本文引用:3][JCR: 5.242]
[17]
PickeringF S, ReisP J. Effects of abomasal supplements of methionine on wool growth of grazing sheep. J Exp Agric, 1993, 33: 7-12[本文引用:1][JCR: 1.621]
[18]
ReisP J. Effects of amino acids on the growth and , 1979. pp223-242[本文引用:1]
[19]
SchroederH E, Khan M R I, KnibbW R, SpencerD, Higgins T J V. Expressing of chicken ovalbumin gene in three Lucerne cultivars. J Plant Physiol, 1991, 18: 495-505[本文引用:1][JCR: 2.699]
[20]
吕德扬, 范云六, 俞梅敏, 唐顺学, 侯宁, 汪清. 苜蓿高含硫氨基酸蛋白转基因植株再生. 遗传学报, 2000, 27: 331-337LüD Y, FanY L, YuM M, TangS X, HouN, WangQ. Regeneration of HNP transgenic alfalfa plants by agrobacterium mediated gene transfer. Acta Genet Sin, 2000, 27: 331-337 (in Chinese with English abstract)[本文引用:1]
[21]
AvrahamT, BadaniH, GaliliS, AmirR. Enhanced levels of methionine and cysteine in transgenic alfalfa (Medicago sativa L. ) plants overexpressing the Arabidopsis cystathionine γ-synthase gene. Plant Biotechnol J, 2005, 3: 71-79[本文引用:1][JCR: 6.279]
[22]
朱延明, 柏锡, 才华, 纪巍, 李勇, 季佐军. 提高大豆蛋氨酸含量的人工序列及其植物表达载体: 中国专利, 编号200910073162. 7. 2010-04-28ZhuY M, BaiX, CaiH, JiW, LiY, JiZ J. Artificial Sequence of Methionine Improved Content in Soybean and Expression Vector Construction. Chinese patent, No. 200910073162. 7 [2010-04-28](in Chinese)[本文引用:1]
[23]
翟红, 柏锡, 朱延明, 陈秀华. SCMRP基因原核表达及多克隆抗体制备. 东北农业大学学报, 2009, 40(7): 60-65ZhaiH, BaiX, ZhuY M, ChenX H. Protokaryotic expression of SCMRP gene and preparation of polyclonal antibody. J Northeast Agric Univ, 2009, 40(7): 60-65 (in Chinese with English abstract)[本文引用:2][CJCR: 0.1053]
[24]
张凤. 转SCMRP基因大豆的遗传稳定性分析及环境安全性初步评价. 东北农业大学硕士学位论文, 2012ZhangF. Field Test and Genetic Stability Analysis of Transgenic SCMRP Soybean Lines. MS Thesis of Northeast Agricultural University, Harbin, China2012 (in Chinese with English abstract)[本文引用:1]
[25]
盛慧, 朱延明, 李杰, 柏锡, 才华. DREB2A基因对苜蓿遗传转化的研究. 草业科学, 2007, 24(3): 40-45ShengH, ZhuY M, LiJ, BaiX, CaiH. Genetic transformation of DREB2A gene into alfalfa. Pratac Sci, 2007, 24(3): 40-45 (in Chinese with English abstract)[本文引用:1]
[26]
陈富成, 祁建民, 徐建堂, 陈涛, 陶爱芬, 林培清, 陈美霞, 郭英, 李华丽. 圆果种黄麻功能叶总蛋白提取方法及双向电泳体系的优化. 作物学报, 2011, 37: 369-373ChenF C, QiJ M, XuJ T, ChenT, TaoA F, LinP Q, ChenM X, GuoY, LiH L. Optimization of extraction method and two- dimensional electrophoresis conditions for proteome analysis of jute functional leaf. Acta Agron Sin, 2011, 37: 369-373 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[27]
SambrookJ, RussellD W. Molecular Cloning: a Laboratory Manual, 3rd edn. New York: Cold Spring Harbor Laboratory Press, 2001. pp1713-1726[本文引用:1]
[28]
HodgesD M, DeLongJ M, ForneyC F, PrangeR K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, 1999, 207: 604-611[本文引用:1][JCR: 3.347]
[29]
HealthR L, PackerL. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys, 1968, 125: 1892-1981[本文引用:1][JCR: 3.37]
[30]
GibonY, LarherF. Cycling assay for nicotinamide adenine dinucleotides: NaCl precipitation and ethanol solubilization of the reduced tetrazolium. Anal Biochem, 1997, 251: 153-157[本文引用:1][JCR: 2.582]
[31]
魏正巍. 耐盐碱GsGST13SCMRP和GsCBRLKSCMRP双价基因对苜蓿的遗传转化及转基因植株的培育. 东北农业大学硕士论文, 2012WeiZ W. The Transformation of Saline-Alkaline Stress Gene into Alfalfa and the Creation of Transgenic Plants. MS Thesis of Northeast Agricultural University, Harbin, China, 2012 (in Chinese with English abstract)[本文引用:1]
[32]
MitsuharaI, UgakiM, HirochikaH, OhshimaM, MurakamiT, GotohY, KatayoseY, NakamuraS, HonkuraR, NishimiyaS, UenoK, MochizukiA, TanimotoH, TsugawaH, OtsukiY, OhshiY. Efficient Promoter cassettes for enhanced expression of foreign genes in dicotyledonous and monocotyledonous plants. Plant Cell Physiol, 1996, 37: 49-59[本文引用:1][JCR: 4.134]
[33]
GarrettR H. Biochemistry 3rd ed. Beijing: Higher Education Press, 2005[本文引用:1]
[34]
杨靓. 野生大豆渗透胁迫相关蛋白激酶基因的克隆及功能分析. 东北农业大学博士学位论文, 2010YangL. Isolation and Functional Analysis of Protein Kinase Genes Relevant to Osmotic Stress in Glycine soja. PhD Dissertation of Northeast Agricultural University, Harbin, China, 2010 (in Chinese with English abstract)[本文引用:1]
[35]
魏正巍, 朱延明, 化烨, 才华, 纪巍, 柏锡, 王臻昱, 文益东. 转GsPPCK1基因苜蓿植株的获得及其耐碱性分析. 作物学报, 2013, 39: 68-75WeiZ W, ZhuY M, HuaY, CaiH, JiW, BaiX, WangZ Y, WenY D. Transgenic alfalfa with GsPPCK1 and its alkaline tolerance analysis. Acta Agron Sin, 2013, 39: 68-75 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[36]
高文俊, 徐静, 谢开云, 董宽虎. Na2CO3和NaHCO3胁迫下冰草的生长及生理响应. 草业学报, 2011, 20(4): 299-304GaoW J, XuJ, XieK Y, DongK H. Physiological responses of Agropyron cristatum under Na2CO3 and NaHCO3 stress. Acta Pratac Sin, 2011, 20(4): 299-304 (in Chinese with English abstract)[本文引用:1]
[37]
李源, 刘贵波, 高洪文, 孙桂枝, 赵海明, 谢楠. 紫花苜蓿种质耐盐性综合评价及盐胁迫下的生理反应. 草业学报, 2010, 19(4): 79-86LiY, LiuG B, GaoH W, SunG Z, ZhaoH M, XieN. A comprehensive evaluation of salt tolerance and the physiological response of Medicago sativa at the seedling stage. Acta Pratac Sin, 2010, 19(4): 79-86 (in Chinese with English abstract)[本文引用:1]
[38]
张海娜, 李小娟, 李存东, 肖凯. 过量表达小麦超氧化物歧化酶(SOD)基因对烟草耐盐能力的影响. 作物学报, 2008, 34: 1403-1408ZhangH N, LiX J, LiC D, XiaoK. Effects of overexpression of wheat superoxide dismutase (SOD) genes on salt tolerant capability in tobacco. Acta Agron Sin, 2008, 34: 1403-1408 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[39]
王玉祥, 张博, 王涛. 盐胁迫对苜蓿叶绿素、甜菜碱含量和细胞膜透性的影响. 草业科学, 2009, 26(3): 53-56WangY X, ZhangB, WangT. Effect of salt stress on the contents of chlorophyll and betaine and its membrane permeability of Medicago sativa. Pratac Sci, 2009, 26(3): 53-56 (in Chinese with English abstract)[本文引用:1]
[40]
李宝健, 朱华晨. 论应用多基因转化策略综合改良生物体遗传性研究方向的前景. 中山大学学报(自然科学版), 2005, 44(4): 79-83LiB J, ZhuH C. On the prospects of applying the multi-gene transformation strategy (MTS) to modify the inheritance of organisms: II. General principles, possible problems and prospects of the MTS. Acta Sci Nat Univ Sunyatseni (Nat Sci Edn), 2005, 44(4): 79-83 (in Chinese with English abstract)[本文引用:1]
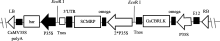


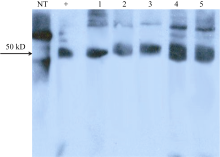



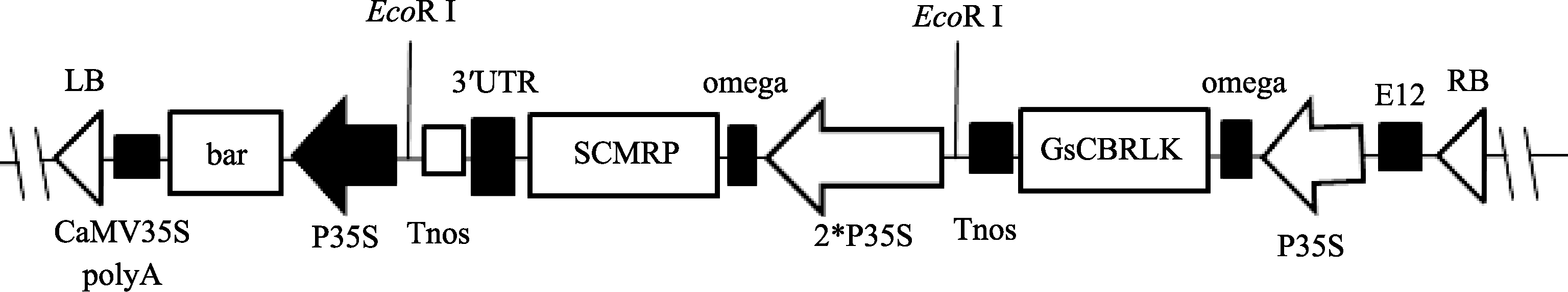






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}