关键词:高丹草; 遗传效应; AFLP; SSR; SRAP; 杂种优势; 预测模型 Genetic Effects and Heterosis Prediction Model ofSorghum bicolor×S. sudanense Grass LU Xiao-Ping1, LIU Dan-Dan1, WANG Shu-Yan1, MI Fu-Gui2, HAN Ping-An1, L#cod#x000dc; Er-Suo1 1 Agronomy College, Inner Mongolia Agricultural University, Huhhot 010019, China
2 College of Ecology and Environmental Science, Inner Mongolia Agricultural University, Huhhot 010019, China
Fund: AbstractMolecular markers-based heterosis prediction can provide some advices forSorghum bicolor×S. sudanense breeding. The 90 cross combinations were made according to the North Carolina design II (NCII) with five sorghum sterile lines as maternal parents and 18 Sudan grass lines as paternal parents. The field trials were carried out on the farms in Hohhot and Baotou to evaluate environmental effects. The parental genetic differences were compared by yield related QTL markers. The phenotypic values of eight traits for all F1 hybrids were investigated. The values were used for selecting significant marker loci on parental lines for setting up the evaluation system of marker effect and marker value. The specific loci were used for evaluating the trait effects and hybrid marker value, and for analyzing the correlation between the marker value and heterosis. The prediction models of the eight traits for the hybrid were constructed with the stepwise regression analysis. The Jackknife sampling method was used to test the accuracy and stability of the model. The result indicated that, considering dominance and additive effect separately, eight traits showed the average correlation coefficient of 0.65 between marker value and phenotypic value. The coefficient of determination was from 0.51 to 0.88 in the eight traits. The results in two places were similar. The model could be instructive for heterosis prediction and parents selection.
Keyword:Sorghum bicolor × S. sudanense; Genetic effect; AFLP; SSR; SRAP; Heterosis; Prediction model Show Figures Show Figures
表1 与高丹草产量相关的QTL标记 Table 1 Yield related QTLs markers on sorghum-sudan grass
引物 Primer
染色体 Chromosome
QTL
参考文献 Reference
引物 Primer
染色体 Chromosome
QTL
参考文献 Reference
E41/M60
6
Fwp6
[21-23]
E45/M60
8
Ph8
[21-23]
E41/M49
4
Fwp4
[21-23]
E41/M48
2
u2a
[21-23]
me8/em9
6
Lw6
[23-24]
C12
3
Ph3.2
[25, 28]
E37/M59
3
Fwp3
[21-23]
me5/em7
1
Sd1
[23-24]
E45/M61
2
Ph2
[21-23]
E39/M51
10
Ph10
[21-23]
E36/M59
7
Ph7
[21-23]
E39/M53
1
tn1
[21-23]
E45/M60
1
sd1
[21-23]
E45/M57
2
tn2
[21-23]
E43/M49
4
sd4
[21-23]
E44/M53
3
tn3
[21-23]
B3235
5
Ph5.2
[25, 28]
E39/M51
1
fwp1a
[21-23]
E45/M60
6
sd6
[21-23]
E40/M47
4
tn4
[21-23]
E37/M52
7
sd7
[21-23]
E32/M49
3
ln3
[21-23]
E43/M49
7
ln7
[21-23]
E43/M49
8
n8
[21-23]
E40/M59
10
n10
[21-23]
D2318
2
Ph2.1
[25, 28]
E34/M53
2
u2b
[21-23]
E41/M60
6
u6
[21-23]
me9/em11
2
Ph2
[23-24]
me3/em4
3
Ln3
[23-24]
E40/M60
9
u9
[21-23]
E41/M57
10
u10
[21-23]
E41/M58
1
w1a
[21-23]
E45/M60
1
w1b
[21-23]
E41/M48
2
w2a
[21-23]
E41/M68
2
lw2b
[21-23]
E37/M59
1
el1b
[21-23]
E48/M60
4
el4
[21-23]
me9/em5
3
Hl3
[23-24]
me5/em4
1
Ll1
[23]
E39/M55
10
el10
[21-23]
E42/M60
10
lw10
[21-23]
表1 与高丹草产量相关的QTL标记 Table 1 Yield related QTLs markers on sorghum-sudan grass
表2 Table 2 表2(Table 2)
表2 供试材料的编号及来源 Table 2 Accession number and sources of experiment materials
编号 Code
名称 Accession
来源 Source
P1
11A
山西省农业科学院 SAAS
P2
13A
内蒙古农业大学农学院 ACIMAU
P3
314A
山西省农业科学院 SAAS
P4
2397A
赤峰市农业科学院高粱研究所 SICAAS
P5
赤12 A Chi 12A
赤峰市农业科学院高粱研究所 SICAAS
P6
Nhei 073
13A×白壳苏丹草后代 13A×White skull Sudan grass progeny
P7
05- P-SD
苏丹草后代 Sudan grass progeny
P8
GZ-40
314A×棕壳苏丹草后代 314A×Brown skull Sudan grass progeny
P9
黑壳苏丹草 Black Skull Sudan grass
苏丹草后代 Sudan grass progeny
P10
白壳苏丹草 White Skull Sudan grass
苏丹草后代 Sudan grass progeny
P11
GZ-186
314A×棕壳苏丹草后代 314A×Brown Skull Sudan grass progeny
P12
GZ21
314A×棕壳苏丹草后代 314A×Brown Skull Sudan grass progeny
P13
GZ-126
2397A×GZ-12
P14
GZ-92
13A×21
P15
GZ-49
13A×GZ-126
P16
GZ-142
13A×GZ-49
P17
Nhei074
2397A×白壳苏丹草 2397A×White Skull Sudan grass
P18
Nhei072
314A×白壳苏丹草后代 314A×White Skull Sudan grass progeny
P19
Nhei071
314A×白壳苏丹草后代 314A×White Skull Sudan grass progeny
P20
62
11A×GZ-92
P21
GZ-12
11A×GZ-142
P22
GZ-183
11A×GZ-12
P23
GZ-190
314A×棕壳苏丹草后代 314A×Brown Skull Sudan grass progeny
SAAS: Shanxi Academy of Agricultural Sciences; ACIMAU: Agronomy College of Inner Mongolia Agricultural University; SAAS: Shanxi Academy of Agricultural Sciences; SICAAS: Sorghum Institute of Chifeng Academy of Agricultural Sciences; SICAAS: Sorghum Institute of Chifeng Academy of Agricultural Sciences.
表2 供试材料的编号及来源 Table 2 Accession number and sources of experiment materials
表6 亲本及子代F1单株鲜重标记型值 Table 6 Marker effect values of plant fresh weight in F1 and their parents
编号 Code
P1
P2
P3
P4
P5
0.0005
0.1375
0.2945
0.4545
0.5238
P6
-0.5445
0.3370
0.2265
-1.2255
0.4825
0.3146
P7
0.0625
-0.5155
-0.0840
0.6920
-0.7930
-0.8245
P8
-0.3885
0.3886
0.2140
-1.0580
-0.5530
-0.7891
P9
0.1105
-0.8380
-0.3895
-0.7565
-0.6685
0.6932
P10
-0.3705
-0.6175
-0.1860
-1.0000
-0.8950
0.2176
P11
0.7825
-1.3495
-0.6830
-0.9410
-0.8930
-1.2953
P12
0.2805
-0.8960
0.1955
-0.8965
-1.0055
0.7321
P13
0.1055
0.1725
0.2590
0.1810
0.4500
0.1085
P14
-0.2865
-0.5630
-0.5635
-0.5155
-0.8405
-0.9354
P15
0.1915
-0.6865
0.2705
-0.7420
-0.8510
1.8897
P16
0.1365
-0.9685
0.3660
-0.9380
1.1330
0.1698
P17
0.0705
-0.4280
0.0035
-0.3505
-0.8915
0.7868
P18
0.3585
0.3615
-0.9140
-1.1180
-0.5031
-0.6191
P19
0.2135
-0.7389
-0.4815
-0.8065
-0.5692
0.7192
P20
-0.4104
-0.5973
-0.2161
-1.1230
0.9052
0.2512
P21
-1.2825
-1.2415
-0.7136
-1.0412
0.9231
-1.5651
P22
0.3201
-0.9564
0.2586
-0.9261
-1.2359
0.9520
P23
0.5158
0.2822
0.2195
0.2616
0.6523
0.4186
表中第2行数据为母本标记型值, 第2列数据为父本标记型值。 The data in the second horizontal line and the second vertical line are the maternal and the paternal marker values, respectively.
表6 亲本及子代F1单株鲜重标记型值 Table 6 Marker effect values of plant fresh weight in F1 and their parents
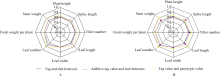
图1 杂种标记型值与性状表型值的相关性A代表呼和浩特的样本数据分析; B代表包头的样本数据分析。Fig. 1 Correlation between marker-type values of hybrid and their phenotypeA: data analysis came from Huhhot sample; B: data analysis came from Baotou sample.
詹秋文, 钱章强. 高粱与苏丹草杂种优势利用的研究. 作物学报, 2004, 30: 73-77ZhanQ W, QianZ Q. Research of sorghum-Sudan grass of heterosis utilization. Acta Agron Sin, 2004, 30: 73-77 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[2]
王和平, 张福耀, 程庆军, 平俊爱, 杜志宏, 郭玉兰, 王晋中. 高粱-苏丹草杂交草的研究与利用. 杂粮作物, 2000, 20(4): 20-23WangH P, ZhangF Y, ChengQ J, PingJ A, DuZ H, GuoY L, WangJ Z. The Research and utilization of sorghum-Sudan grass. Rain Fed Crops, 2000, 20(4): 20-23 (in Chinese)[本文引用:1][CJCR: 0.5562]
[3]
赵甘霖, 丁国祥, 熊洪, 曾庆曦. 我国高粱-苏丹草杂交种研究进展及应用前景. 西南农业学报, 2003, 16(4): 126-129ZhaoG L, DingG X, XiongH, ZengQ X. Prospect of progress and application of sorghum-Sudan grass in China. Southwest China J Agric Sci, 2003, 16(4): 126-129 (in Chinese with English abstract)[本文引用:1]
[4]
ChungJ M, LeeB C, KimJ S. Fine-scale structure among genetic individuals of clone-forming monotypic genus Echinosophora koreensis (Fabaceae). Ann Bot, 2006, 98: 165-173[本文引用:1][JCR: 0.657]
[5]
钱章强. 利用高粱雄性不育系选育饲用新品种. 中国草地, 1990, (6): 61-71QianZ Q. Breeding of new variety with male sterile line of sorghum. China Grassland , 1990, (6): 61-71 (in Chinese with English abstract)[本文引用:1]
[6]
牟芝兰. 刈割周期对苏丹草叶片生长的影响. 草业科学, 1994, 11(6): 29-31MouZ L. Research of foliage growth of sorghum-Sudan grass[本文引用:1]
[7]
aftercutting. Pratac Sci, 1994, 11(6): 29-31 (in Chinese with English abstract)[本文引用:1]
[8]
陈才夫, 王槐三. 苏丹草×拟高粱种间杂种主要特性及细胞学分析. 南京农业大学学报, 1993, (2): 58-63ChenC F, WangH S. Main characteristics and cytological analysis of Sudan grass × sorghum interspecific hybrids. J Nanjing Agric Univ, 1993, (2): 58-63 (in Chinese with English abstract)[本文引用:1][CJCR: 0.916]
[9]
孟钢. “先锋”高丹草——饲用高粱和苏丹草杂交品种. 世界农业, 2003, (6): 46-48MengG. “Pioneer” sorghum-Sudan grass ― hybrid of forage sorghum and Sudan grass. World Agric, 2003, (6): 46-48 (in Chinese)[本文引用:1][CJCR: 0.712]
[10]
于卓, 赵晓杰, 秦永梅. 高粱与苏丹草2个杂交组合F2代的生育及细胞遗传学研究. 草业科学, 2004, 7: 13-17YuZ, ZhaoX J, QinY M. Growth and cyto-genetics of F2 generation in Sorghum × Sudan grass combinations. Pratac Sci, 2004, 7: 13-17 (in Chinese with English abstract)[本文引用:1]
[11]
赵晓杰, 于卓, 刘永伟, 逯晓萍. 高粱314A、13A与苏丹草杂种F1代的农艺特性及细胞学分析. 西北植物学报, 2005, 25: 1107-1113ZhaoX J, YuZ, LiuY W, LuX P. Agronomic trait and cytology of hybrid F1 of sorghum (314A and 13A) × Sudan grass. Acta Bot Boreali-Occident Sin, 2005, 25: 1107-1113 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[12]
于卓, 秦永梅, 赵晓杰, 赵娜, 闫文芝. 优良饲用作物新品种——蒙农青饲1号高丹草选育. 中国草地, 2004, 26(2): 1-9YuZ, QinY M, ZhaoX J, ZhaoN, YanW Z. New forage variety― sorghum × Sudan grass of Mengnongqingsi 1# breeding. China Grassland , 2004, 26(2): 1-9 (in Chinese with English abstract)[本文引用:1]
[13]
于卓, 赵晓杰, 赵娜, 秦永梅, 刘永伟. 蒙农青饲2号高丹草选育. 草地学报, 2004, 12: 176-182YuZ, ZhaoX J, ZhaoN, QinY M, LiuY W. Sorghum × sudan grass of Mengnongqingsi 2# breeding. Acta Agres Sin, 2004, 12: 176-182 (in Chinese with English abstract)[本文引用:1]
[14]
于卓, 马艳红, 李小雷, 刘志华, 周亚星, 李晓宇, 李长青. 蒙农3号高丹草选育. 中国草地学报, 2008, 30(6): 1-9YuZ, MaY H, LiX L, LiuZ H, ZhouY X, LiX Y, LiC Q. Sorghum × Sudan grass of Mengnongqingsi 3# breeding. Chin J Grassland , 2008, 30(6): 1-9 (in Chinese with English abstract)[本文引用:1][CJCR: 1.018]
[15]
詹秋文, 林平, 钱章强. 皖草3号的选育及其特征特性. 作物杂志, 2006, (4): 35-36ZhanQ W, LinP, QianZ Q. The characteristic and breeding of Wancao 3#. Crops, 2006, (4): 35-36 (in Chinese)[本文引用:1][CJCR: 0.6276]
[16]
赵庆勇, 朱镇, 张亚东, 赵凌, 陈涛, 张巧凤, 王才林. SSR标记遗传距离与粳稻杂种优势的相关性分析. 中国水稻科学, 2009, 27: 141-147ZhaoQ Y, ZhuZ, ZhangY D, ZhaoL, ChenT, ZhangQ F, WangC L. Correlation analysis between genetic distances based SSR and heterosis of japonica rice. Chin J Rice Sci, 2009, 27: 141-147 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[17]
张培江, 才宏伟, 李焕朝, 杨联松, 张德泉, 白一松, 胡兴明, 许传万. RAPD分子标记水稻遗传距离及其与杂种优势的关系. 安徽农业科学, 2000, 28: 697-700ZhangP J, CaiH W, LiH C, YangL S, ZhangD Q, BaiY S, HuX M, XuC W. Relationship between genetic distance and heterosis of rice based on RAPD marker. J Anhui Agric Sci, 2000, 28: 697-700 (in Chinese with English abstract)[本文引用:1][CJCR: 0.687]
[18]
张涛, 韩磊, 徐建第, 蒋开锋, 吴先军, 汪旭东, 郑家奎. 杂交香稻亲本遗传距离与产量杂种优势的相关性研究. 中国农业科学, 2006, 39: 831-835ZhangT, HanL, XuJ D, JiangK F, WuX J, WangX D, ZhengJ K. Correlation analysis between genetic distance and yield heterosis of scented rice. Sci Agric Sin, 2006, 39: 831-835 (in Chinese with English abstract)[本文引用:1]
[19]
袁力行, 傅骏骅, 刘新芝, 彭泽斌, 张世煌, 李新海, 李连城. 利用分子标记预测玉米杂种优势的研究. 中国农业科学, 2000, 33(6): 6-12YuanL X, FuJ H, LiuX Z, PengZ B, ZhangS H, LiX H, LiL C. Prediction of heterosis of maize based on molecular marker. Sci Agric Sin, 2000, 33(6): 6-12 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[20]
ZhangQ F, GaoY J, Saghui Marrof M A, et al. Molecular divergence and hybrid performance in rice. Mol Breed, 1995, 1: 133-142[本文引用:1][JCR: 3.251]
[21]
张涛, 倪先林, 蒋开锋, 杨乾华, 万先齐, 曹应江. 水稻功能基因标记遗传距离与杂种优势的相关性研究. 中国水稻科学, 2009, 23: 567-572ZhangT, NiX L, JiangK F, YangQ H, WanX Q, CaoY J. Correlation between genetic distance based on molecular markers of functional genes and heterosis in rice. Chin J Rice Sci, 2009, 23: 567-572 (in Chinese with English abstract)[本文引用:4][CJCR: 1.494]
[22]
逯晓萍, 云锦凤, 肖宇红, 米福贵, 李美娜, 尹利. 高丹草(高粱×苏丹草)产量及其构成因素的QTL定位与分析. 华北农学报, 2007, 22(4): 80-85LuX P, YunJ F, XiaoY H, MiF G, LiM N, YinL. QTLs of yield related traits on sorghum-Sudan grass. Acta Agric Boreali-Sin, 2007, 22(4): 80-85 (in Chinese with English abstract)[本文引用:3][CJCR: 0.951]
[23]
逯晓萍, 云锦凤. 高丹草遗传图谱构建及农艺性状基因定位研究. 草地学报, 2005, 13: 262-263LuX P, YunJ F. Research on genetic mapping and agronomic gene location of sorghum-Sudan grass. Acta Agres Sin, 2005, 13: 262-263 (in Chinese with English abstract)[本文引用:3]
[24]
LuX P, YunJ F, GaoC P, SuryaA. QTL Analysis of economically important traits in Sorghum bicolor × S. sudanense hybrid. Can J Plant Sci, 2011, 91: 81-90[本文引用:3][JCR: 0.716]
[25]
刘丹丹, 逯晓萍, 张瑞霞, 温莹, 薛春雷, 杨凯, 任瑞, 王亚男, 韩平安. 高丹草种质资源SRAP指纹图谱构建及遗传多样性分析. 华北农学报, 2012, 27(2): 1-8LiuD D, LuX P, ZhangR X, WenY, XueC L, YangK, RenR, WangY N, HanP A. SRAP fingerprint map and genetic diversity of sorghum-Sudan germplasm. Acta Agric Boreali-Sin, 2012, 27(2): 1-8 (in Chinese with English abstract)[本文引用:3][CJCR: 0.951]
[26]
温莹, 逯晓萍, 任锐, 米福贵, 韩平安, 薛春雷. 高丹草EST-SSR标记的开发及其遗传多样性. 遗传, 2013, 35: 225-232WenY, LuX P, RenR, MiF G, HanP A, XueC L. Development of EST-SSR marker and genetic diversity analysis in Sorghum bicolor × S. sudanenes. Hereditas (Beijing), 2013, 35: 225-232 (in Chinese with English abstract)[本文引用:1]
[27]
逯晓萍, 云锦凤, 张雅慧, 薛春雷, 陈强. 高丹草重组自交系群体的遗传变异与高产种质的创新. 华北农学报, 2009, 24(5): 90-95LuX P, YunJ F, ZhangY H, XueC L, ChenQ. Genetic variation of inbred population and creation of high yield germ on Sorghum bicolor × S. sudanense hybrid. Acta Agric Boreali-Sin, 2009, 24(5): 90-95 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
[28]
逯晓萍, 云锦凤, 米福贵, 陈强, 张雅慧, 薛春雷. 基于性状和分子标记的高丹草近等基因系的分离研究. 中国农业科学, 2010, 43: 468-473LuX P, YunJ F, MiF G, ChenQ, ZhangY H, XueC L. Segregation of near isogene lines of Sorghum bicolor × S. sudanense based on traits and molecular marker. Sci Agric Sin, 2010, 43: 468-473 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[29]
苏舒, 董维, 游录鹏, 黄守程, 戚金亮, 陆桂华, 黄应华, 杨永华. 高粱株高性状的QTL定位初步分析. 江苏农业科学, 2012, 40(3): 19-21SuS, DongW, YouL P, HuangS C, QiJ L, LuG H, HuangY H, YangY H. Primary analysis of QTLs location related height of sorghum. Jiangsu Agric Sci, 2012, 40(3): 19-21 (in Chinese with English abstract)[本文引用:1]
[30]
张南松, 陈军杰, 祝增荣. 利用多重抽样估计稀有物种的种类数. 生物数学学报, 2002, 17: 455-460ZhangN S, ChenJ J, ZhuZ R. Multiple sampling evaluation of the number of rare species. J Math Biol, 2002, 17: 455-460 (in Chinese with English abstract)[本文引用:1][JCR: 2.366]
[31]
许国. Jackknife估计及其应用浅探. 山东师范大学学报, 2000, 15: 454-455XuG. Jackknife evaluation and application. J Shand ong Normal Univ, 2000, 15: 454-455 (in Chinese with English abstract)[本文引用:1]
[32]
TamsS H, BauerE, OettlerG. Prospects for hybrid breeding in winter triticale: II. Relationship between parental genetic distance and specific combining ability. Plant Breed, 2006, 125: 331-336[本文引用:1][JCR: 1.175]
[33]
HariD, Upadhyaya, WangY H. SSR markers linked to kernel weight and tiller number in sorghum identified by association mapping. Euphytica, 2012, 187: 401-410[本文引用:1][JCR: 1.643]
[34]
何光华, 侯磊, 李德谋, 罗小英, 牛国清, 唐梅, 裴炎. 利用分子标记预测杂交水稻产量及其构成因素. 遗传学报, 2002, 29: 438-444HeG H, HouL, LiD M, LuoX Y, NiuG Q, TangM, PeiY. Prediction of yield and related trait factors of hybrid rice with molecular marker. Acta Genet Sin, 2002, 29: 438-444 (in Chinese with English abstract)[本文引用:1]
[35]
游书梅, 曹应江, 郑家奎, 蒋开锋, 张涛, 杨莉, 杨乾华, 万先齐, 罗婧. 杂交籼稻子粒QTL标记遗传距离与粒重、粒重杂种优势的相关性. 植物遗传资源学报, 2012, 13: 1050-1054YouS M, CaoY J, ZhengJ K, JiangK F, ZhangT, YangL, YangQ H, WanX Q, LuoJ. Correlation of genetic distance and seed weight heterosis based on QTL markers of hybrid rice. J Plant Genet Resour, 2012, 13: 1050-1054 (in Chinese with English abstract)[本文引用:1][CJCR: 1.1628]
[36]
武耀廷, 张天真, 朱协飞, 王广明. 陆地棉遗传距离与杂种F1、F2产量及杂种优势的相关分析. 中国农业科学, 2002, 35: 22-28WuY T, ZhangT Z, ZhuX F, WangG M. Correlation of genetic distance, yield of F1 and F2 and hybrid heterosis. Sci Agric Sin, 2002, 35: 22-28 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[37]
VuylstekeM, KuiperM, StamP. Chromosomal regions involved in hybrid performance and heterosis: their AFLP(R)-based identification and practical use in prediction models. Heredity, 2000, 85: 208-218[本文引用:1][JCR: 4.11]

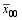

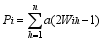
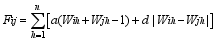
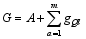



{kind=link}