关键词:蓖麻; 株高性状; QTL定位; 遗传解析 Genetic Analysis of Traits Related to Plant Height inRicinus communisL. Based on QTL Mapping LIU Chen**, LU Jian-Nong**, YIN Xue-Gui*, BI Chuan, WEN Dan-You, ZHENG Jun, LIU Shuai, SHI Zhuo-Xing, CHENG Yue-Xiang Agricultural College of GuangDong Ocean University, Zhanjiang 524088, China Fund: AbstractThe traits related to plant height were analyzed with correlation, regression and QTL mapping in two F2 populations derived from crosses of YC2×YF1 and YC1×YF1, respectively. The results indicated that plant height (PH) was significantly and positively correlated with bearing height of primary raceme (PRH), length of main stem internode (MSIL) and main stem diameter (MSD) but not with node number of main stem (MSNN), PRH was significantly and positively correlated with MSNN, MSIL and MSD, MSNN and MSIL was significantly and negatively correlated with each other. In F2 population from a cross of YC2×YF1, five, four, six, three, and two QTLs for PH, PRH, MSNN, MSIL, and MSD were detected by QTLnetwork 2.0, which explained for 45.9%, 45.3%, 66.1%, 55.4%, and 12.6% of the phenotypic variation, respectively. In F2 population from a cross of YC1×YF1, three, four, five, one, and two QTLs for the five traits mentioned above were detected, which explained for 45.9%, 45.3%, 66.1%, 55.4%, and 12.6% of the phenotypic variation, respectively. We concluded that the pleiotropy or linkage between QTLs results in the positive correlation between PH, PRH and MSIL, the additive effect was the main genetic component of PH, PRH, MSIL, and the epistatic effect is the main genetic component of MSNN and MSD. It suggested that PRH and MSIL should be used to select and predict PH indirectly in the early stage, more MSNN and shorter MSIL should be regarded as the target traits for high-yield breeding.
Keyword:Ricinus communis L.; Traits related to plant height; QTL mapping; Genetic analysis Show Figures Show Figures
表2 性状间的相关系数 Table 2 Correlation coefficients between traits
性状 Trait
株高 PH
主穗位高 PRH
主茎节数 MSNN
主茎节长 MSIL
主穗位高 PRH
0.658**
主茎节数 SNN
-0.054
0.349**
主茎节长 MSIL
0.734**
0.775**
-0.397**
茎粗 MSD
0.579**
0.474**
0.235**
0.347**
**表示显著水平为0.01。 ** Significance at 0.01 probability level. PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode; MSD: main stem diameter.
表2 性状间的相关系数 Table 2 Correlation coefficients between traits
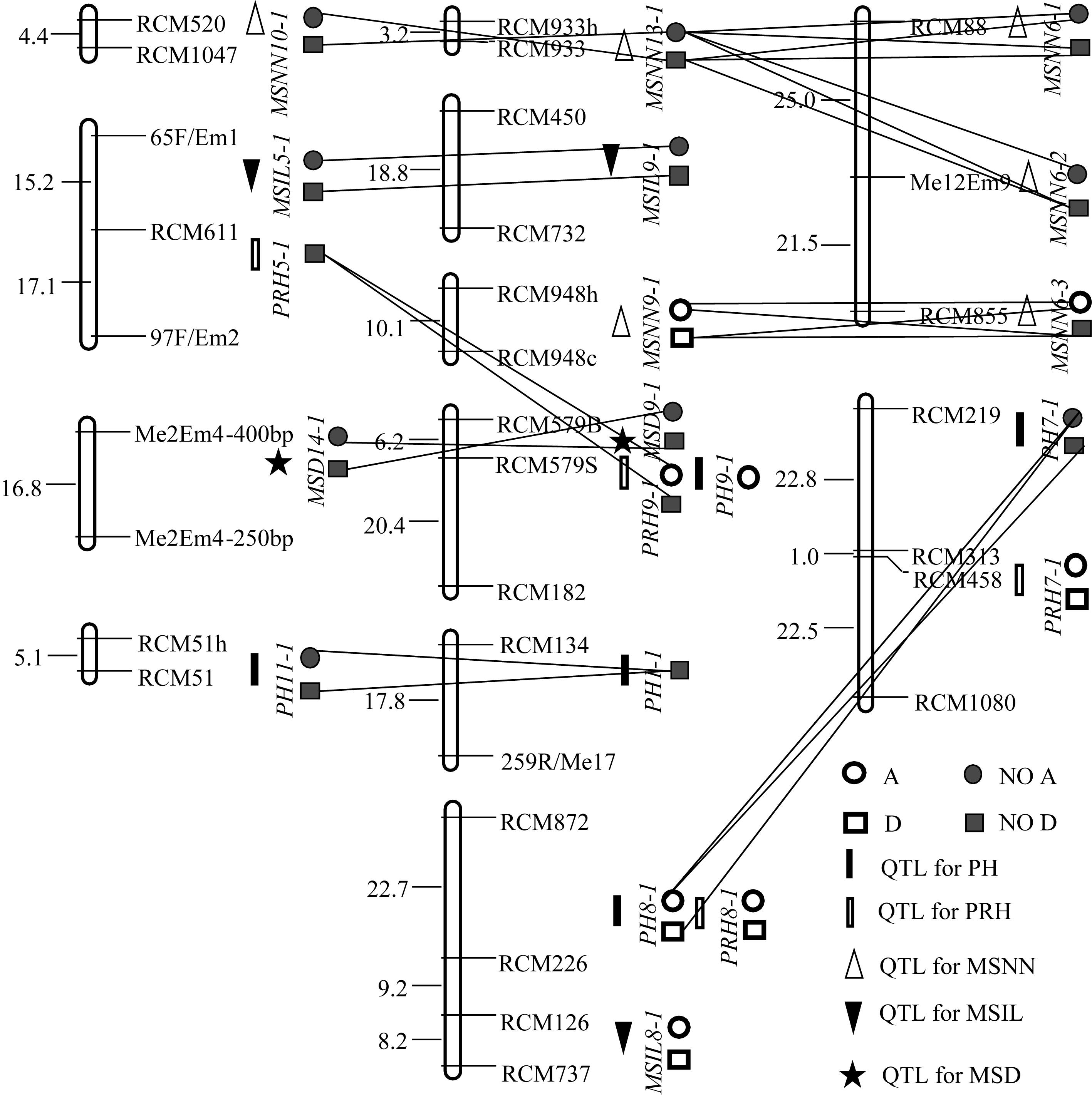
表3 (YC2×YF1)F2群体的QTL定位结果 Table 3 QTLs detected in (YC2×YF1)F2 population
性状 Trait
QTL
标记区间 Marker interval
标记位置 Marker position
QTL位置 QTL position
置信区间 Confidence interval
株高 PH
PH1-1
RCM134-259R/Me17
26.0-43.8
41.0
33.0-47.8
PH7-1
RCM219-RCM313
0-22.8
1.0
0-18.0
PH8-1
RCM872-RCM226
33.0-55.7
46.0
31.0-48.0
PH9-1
RCM579S-RCM182
242.7-263.1
242.7
237.5-250.7
PH11-1
RCM51h-RCM51
0-5.1
5.0
4.0-5.0
主穗位高 PRH
PRH5-1
RCM611-97F/Em2
330.5-347.6
334.5
326.3-341.5
PRH7-1
RCM458-RCM1080
23.8-46.3
23.8
19.0-33.8
PRH8-1
RCM872-RCM226
33.0-55.7
46.0
20.0-48.0
PRH9-1
RCM579S-RCM182
242.7-263.1
242.7
238.5-253.7
主茎节数 MSNN
MSNN6-1
RCM88-Me12Em9
91.1-116.1
91.1
91.1-98.1
MSNN6-2
Me12Em9-RCM855
116.1-137.6
116.1
115.1-121.1
MSNN6-3
RCM855-ISSR836
137.6-137.6
137.6
132.1-141.6
MSNN9-1
RCM948h-RCM948c
174.0-184.1
178.0
167.2-182.0
MSNN10-1
RCM520-RCM1047
43.8-48.2
43.8
38.8-45.8
MSNN13-1
RCM933h-RCM933
0-3.2
3.0
1.0-8.2
节间长 MSIL
MSIL5-1
65F/Em1-RCM611
315.3-330.5
320.3
315.3-327.3
MSIL8-1
RCM126-RCM737
64.9-73.1
65.9
64.9-73.7
MSIL9-1
RCM450-RCM732
74.8-93.6
81.8
74.8-91.8
茎粗 MSD
MMSD9-1
RCM579B-RCM579S
236.5-242.7
241.5
235.5-242.5
MMSD14-1
Me2Em4-250bp-Me2Em4-400bp
0-16.8
16.0
0-16.0
PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode; MSD: main stem diameter.
表3 (YC2×YF1)F2群体的QTL定位结果 Table 3 QTLs detected in (YC2×YF1)F2 population
表4 Table 4 表4(Table 4)
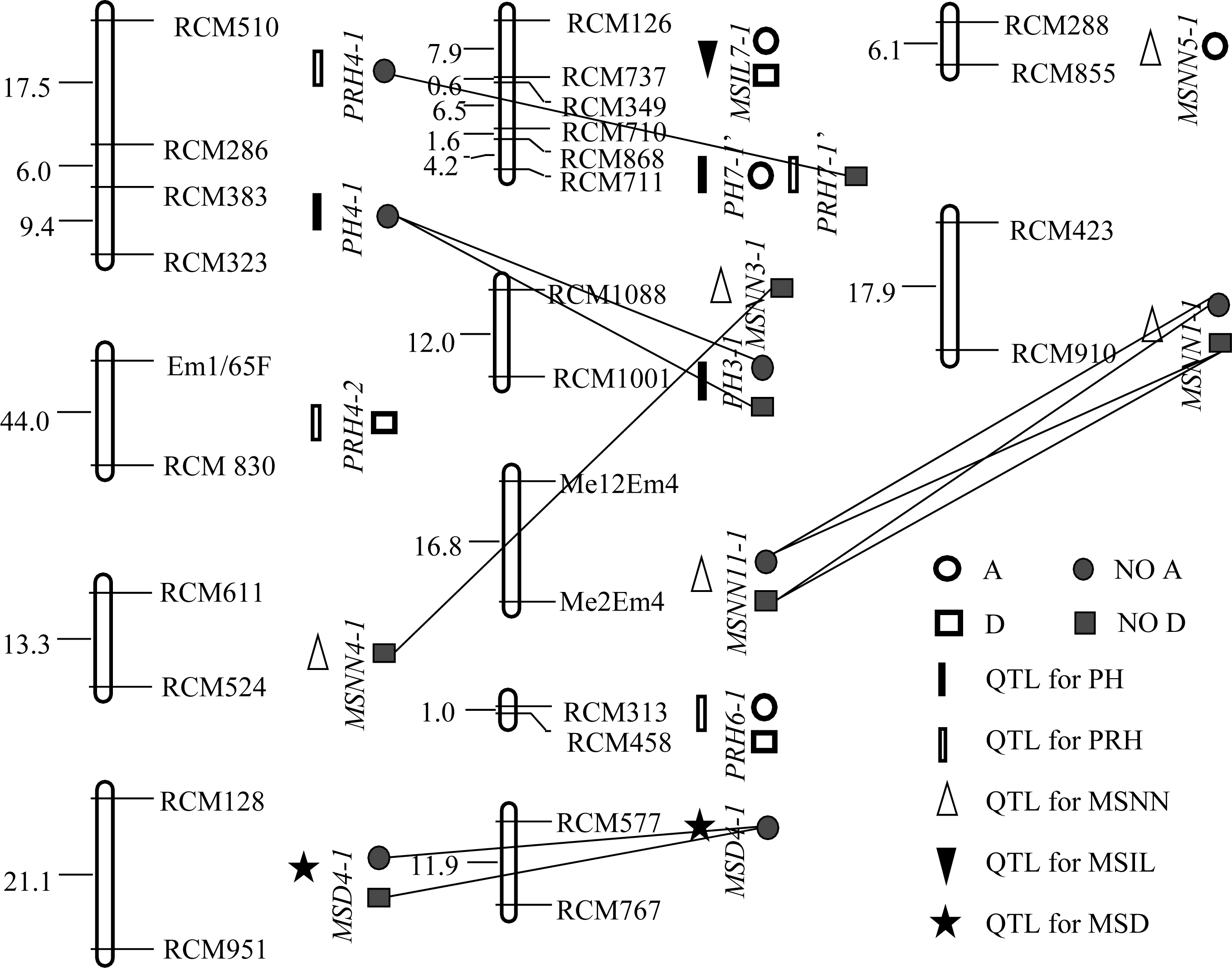
表4 (YC1×YF1)F2群体的QTL定位结果 Table 4 QTLs detected in (YC1×YF1)F2 population
性状 Trait
QTL
标记区间 Marker interval
标记位置 Marker position
QTL位置 QTL position
置信区间 Confidence interval
株高 PH
PH3-1
RCM1088-RCM1001
9.8-21.8
17.8
10.8-21.8
PH4-1
RCM383-RCM323
48.8-58.2
48.8
45.8-51.8
PH7-1’
RCM868-RCM711
38.1-42.3
41.1
38.1-42.1
主茎穗位高 PRH
PRH4-1
RCM510-RCM286
25.3-42.8
31.3
25.3-40.3
PRH4-2
65F/Em1-RCM830
143.5-187.5
166.5
155.5-189.5
PRH6-1
RCM313-RCM458
0-1.0
0
0
PRH7-1’
RCM868-RCM711
38.1-42.1
39.1
30.0-42.1
主茎节数 MSNN
MSNN1-1
RCM423-RCM910
44.9-62.8
61.9
53.9-61.9
MSNN3-1
RCM1088-RCM1001
9.8-21.8
9.8
8.5-13.8
MSNN4-1
RCM611-RCM524
230.2-243.5
230.2
230.2-237.2
MSNN5-1
RCM288-RCM855
53.8-59.9
56.8
43.1-64.9
MSNN11-1
Me12Em4-Me2Em4
0-16.8
16.0
8.0-16.0
节间长 MSIL
MSIL7-1
RCM126-RCM737
21.5-29.4
28.5
25.5-32.0
茎粗 MSD
MSD4-1
RCM128-RCM951
467.1-488.2
483.1
474.1-509.2
MSD10-1
RCM577-RCM767
16.0-27.9
16.0
11.4-21.0
PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode; MSD: main stem diameter.
表4 (YC1×YF1)F2群体的QTL定位结果 Table 4 QTLs detected in (YC1×YF1)F2 population
表5 Table 5 表5(Table 5)
表5 (YC2×YF1)F2群体的QTL遗传效应 Table 5 QTL effects in (YC2×YF1)F2 population
性状 Trait
QTLi
QTLj
ai ( h2a)
di ( h2d)
aj ( h2a)
dj ( h2d)
aaij ( h2aa)
adij ( h2ad)
daij ( h2da)
ddij ( h2dd)
株高 PH
PH9-1
-10.34*** (6.04%)
PH7-1
PH8-1
33.59*** (23.7%)
20.71*** (2.6%)
17.8*** (0.7%)
26.5*** (3.23%)
-19.7*** (2.1%)
PH1-1
PH11-1
-40.7*** (1.6%)
107.0*** (5.9%)
主穗位高 PRH
PRH7-1
PRH8-1
8.01*** (8.0%)
14.88*** (7.4%)
13.0*** (11.6%)
9.28*** (3.0%)
PRH5-1
PRH9-1
-8.92*** (4.9%)
-17.5*** (5.5%)
-26.0*** (4.9%)
主茎节数 MSNN
MSNN6-3
MSNN9-1
-2.02*** (9.0%)
2.04*** (9.6%)
-1.6*** (0.8%)
-4.5*** (14.0%)
4.7*** (6.6%)
-3.8*** (3.76%)
4.9*** (3.07%)
MSNN6-1
MSN13-1
2.3*** (4.5%)
-3.3*** (0.35%)
4.4*** (0.01%)
3.9*** (0.01%)
MSNN6-2
MSN13-1
-8.07*** (0.32%)
-9.14*** (5.1%)
-5.85*** (1.81%)
MSN10-1
MSN13-1
1.38*** (0.16%)
-2.8*** (7.0%)
节间长度 MSIL
MSIL8-1
1.33*** (36.28%)
1.07*** (10.4%)
MSIL5-1
MSIL9-1
-1.2*** (6.28%)
2.0*** (2.4%)
茎粗 MSD
MSD9-1
MSD14-1
8.04*** (1.3%)
6.80*** (11.3%)
***表示0.005显著性水平。 ***Significance at 0.005 probability level. PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode; MSD: main stem diameter.
表5 (YC2×YF1)F2群体的QTL遗传效应 Table 5 QTL effects in (YC2×YF1)F2 population
表6 (YC1×YF1)F2群体的QTL遗传效应 Table 6 QTL effects in (YC1×YF1)F2 population
性状 Trait
QTLi
QTLj
ai ( h2a)
di ( h2d)
aj ( h2a)
dj ( h2d)
aaij ( h2aa)
adij ( h2ad)
daij ( h2da)
ddij ( h2dd)
株高 PH
PH7-1’
—
19.95*** (18.14%)
PH3-1
PH4-1
16.95*** (3.96%)
-22.25*** (3.85%)
主穗位高 PRH
PRH4-2
PRH6-1
27.13*** (4.05%)
7.62*** (6.63%)
12.76*** (7.14%)
PRH4-1
PRH7-1’
-22.59*** (7.66%)
主茎节数 MSNN
MSNN5-1
—
-1.04*** (8.77%)
MSNN1-1
MSNN11-1
-1.86*** (6.36%)
-4.98*** (4.09%)
2.37*** (4.67%)
6.10*** (3.14%)
MSNN3-1
MSNN4-1
-4.09*** (8.06%)
主茎节长 MSIL
MSIL7-1
—
1.31*** (36.30%)
1.01*** (1.05%)
茎粗 MSD
MSD4-1
MSD10-1
-5.11*** (3.26%)
6.74*** (3.32%)
6.10*** (1.06%)
***表示0.005显著性水平。 *** Significance at 0.005 probability level. PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode; MSD: main stem diameter
表6 (YC1×YF1)F2群体的QTL遗传效应 Table 6 QTL effects in (YC1×YF1)F2 population
图1 (YC2×YF1)F2群体QTL的分布及互作A、D、NO A和NO D分别表示加性效应、显性效应、无加性效应和无显性效应; 直线相连表示2个QTL的上位性效应。Fig. 1 Distribution and interaction of QTLs in (YC2×YF1)F2 populationA, D, NO A, and NO D refer to additive effect, dominant effect, no additive effect and no dominant effect, respectively. Line joining two QTLs refers to epistatic effect.
MonnaL, KitazawaN, YoshinoR, SuzukiJ, MasudaH, MaeharaY, TanjiM, SatoM, NasuS, MinobeY. Positional cloning of rice semidwarfing gene, sd-1: rice “green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res, 2002, 9: 11-17[本文引用:1][JCR: 4.425]
[2]
PengJ, RichardsD E, HartleyN M, MunrphyG P, DevosK M, FlinthamJ E, BealesJ, FishL J, Worland A J, PelicaF, SudhakarD, ChristouSnape J, GaleM D, HarberdN P. “Green Revolution” gene encode mutant bibberelin response modnlators. Nature, 1999, 400: 256-261[本文引用:1][JCR: 38.597]
[3]
杨松杰, 张晓科, 何中虎, 夏先春, 周阳. 用STS标记检测矮秆基因Rht-B1b和Rht-D1b在中国小麦中的分布. 中国农业科学, 2006, 39: 1680-1688YangS J, ZhangX K, HeZ H, XiaX C, ZhouY. Distribution of dwarfing genes Rht-B1b and Rht-D1b in Chinese bread wheats detected by STS marker. Sci Agric Sin, 2006, 39: 1680-1688 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[4]
嵇怡, 缪旻珉, 陈学好. 植物矮生性状的分子遗传研究进展. 分子植物育种, 2006, 4: 753-771JiY, MiaoM M, ChenX H. Progresses on the molecular genetics of dwarf character in plants. Mol Plant Breed, 2006, 4: 753-771 (in Chinese with English abstract)[本文引用:1][CJCR: 1.23]
[5]
李卓坤, 谢全刚, 朱占玲, 刘金良, 韩淑晓, 田宾, 袁倩倩, 田纪春. 基于QTL定位分析小麦株高的杂种优势. 作物学报, 2010, 36: 771-778LiZ K, XieQ G, ZhuZ L, LiuJ L, HanS X, TianB, YuanQ Q, TianJ C. Analysis of plant height heterosis based on QTL mapping in wheat. Acta Agron Sin, 2010, 36: 771-778 (in Chinese with English abstract)[本文引用:3][CJCR: 1.667]
[6]
杨芳萍, 夏先春, 张勇, 张晓科, 刘建军, 唐建伟, 杨学明, 张俊儒, 刘茜, 李式昭, 何中虎. 光周期和矮秆基因在不同国家小麦品种中的分布及其效应. 作物学报, 2012, 38: 1155-1166YangF P, XiaX C, ZhangY, ZhangX K, LiuJ J, TangJ W, YangX M, ZhangJ R, LiuQ, LiS Z, HeZ H. Distribution of allelic variation for vernalization, photoperiod, and dwarfing genes and their effects on growth period and plant height among cultivars from major wheat producing countries. Acta Agron Sin, 2012, 38: 1155-1166 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[7]
高奋明, 姜勇, 孔德伟, 李仕贵. 水稻株高的遗传控制及其在育种上的应用. 分子植物育种, 2005, 3: 87-93GaoF M, JiangY, KongD W, LiS G. Genetic control of plant height and its utilization in rice. Mol Plant Breed, 2005, 3: 87-93 (in Chinese with English abstract)[本文引用:1][CJCR: 1.23]
[8]
邢永忠, 徐才国, 华金平, 谈移芳, 孙新立. 水稻株高和抽穗期基因的定位和分离. 植物学报, 2001, 43: 721-726XingY Z, XuC G, HuaJ P, TanY F, SunX L. Mapping and isolation of quantitative trait loci controlling plant height and heading date in rice. Acta Bot Sin, 2001, 43: 721-726 (in Chinese with English abstract)[本文引用:1][JCR: 0.599]
[9]
姜树坤, 黄成, 徐正进, 陈温福. 粳稻株高QTL与赤霉素和油菜素内酯合成及信号转导基因相关分析. 中国农业科学, 2010, 43: 2829-2938JiangS K, HuangC, XuZ J, ChenW F. Relationship between QTLs for plant height and its components and genes controlling gibberellin and brassinosteroid biosynthesis and their transduction in japonica rice (Oryza sativa L. ). Sci Agric Sin, 2010, 43: 2829-2938 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[10]
LavanyaC, GopinathV. Inheritance studies for morphological characters and sex expression in pistillate lines of castor (Ricinus communis L. ). Indian J Genet Plant Breed, 2008, 68: 275-282[本文引用:2][JCR: 0.198]
[11]
李金琴, 朱国立, 何智彪, 张智勇, 贾娟霞, 乔文杰, 李靖霞. 蓖麻矮秆性状基因遗传规律研究. 内蒙古农业科技, 2010, (1): 54-56LiJ Q, ZhuG L, HeZ B, ZhangZ Y, JiaJ X, QiaoW J, LiJ X. Study on the genetic regularity of castor bean dwarf characters gene. J Inner Mongolia Agric Sci Tech, 2010, (1): 54-56 (in Chinese with English abstract)[本文引用:2]
[12]
刘鹏. 蓖麻细胞色素P450基因RNAi植物表达载体构建及遗传转化的研究. 沈阳农业大学博士学位论文, 2012LiuP. The Studies on Construction and Genetic Transformation of RNAi Plant Expression Rector Carrying Cytochrome P450 from Castor. PhD Dissertation of Shenyang Agricultural University, Shenyang, China, 2012 (in Chinese with English abstract)[本文引用:1]
[13]
严兴初. 蓖麻种质资源描述规范和数据标准. 北京: 中国农业出版社. 2005. pp12-19YanX C. Descriptions and data stand ard for castor (Ricinus communis L. ). Beijing: China Agriculture Press, 2005. pp12-19(in Chinese)[本文引用:1]
[14]
Land erE S, GreenP. Mapmaker: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics, 1987, 1: 174-181[本文引用:1][JCR: 3.01]
[15]
YangJ, HuC C, HuH, YuR D, XiaZ, YeX Z, ZhuJ. QTLNetwork: mapping and visualizing genetic architecture of complex traits in experimental populations. Bioinformatics, 2008, 24: 721-723[本文引用:1][JCR: 5.323]
[16]
赵彦宏, 朱军, 徐海明, 杨剑, 高用明, 宋佑胜, 石春海, 邢永忠. 基于QTL定位的水稻有效穗数杂种优势预测. 中国水稻科学, 2007, 21: 350-354ZhaoY H, ZhuJ, XuH M, YangJ, GaoY M, SongY S, ShiC H, XingY Z. Predicting heterosis of effective panicle number per plant based on QTL mapping in rice. Chin J Rice Sci, 2007, 21: 350-354 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[17]
高用明, 朱军, 宋佑胜, 何慈信, 石春海, 邢永忠. 水稻永久F2群体抽穗期QTL的上位性及其与环境互作效应的分析. 作物学报, 2004, 30: 849-854GaoY M, ZhuJ, SongY S, HeC X, ShiC H, XingY Z. Use of Permanent F2 population to analyze epistasis and their Interaction effects with environments for QTLs controlling heading date in rice. Acta Agron Sin, 2004, 30: 849-854 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[18]
张志勇, 黄育民, 张凯, 王侯聪, 江良荣. 水稻株高QTL定位及精确性分析. 厦门大学学报(自然科学版), 2008, 47(1): 116-121ZhangZ Y, HuangY M, ZhangK, WangH C, JiangL R. Detection of QTL for plant height in rice (Oryza sativa L. ) and analysis of QTL mapping accuracy. J Xiamen Univ (Nat Sci), 2008, 47(1): 116-121 (in Chinese with English abstract)[本文引用:1]
[19]
王岩, 李卓坤, 田纪春. 利用永久 F2 群体定位小麦株高的QTL. 作物学报, 2009, 35: 1038-1043WangY, LiZ K, TianJ C. Detection of quantitative trait loci for plant height using an immortalized F2 population in wheat. Acta Agron Sin, 2009, 35: 1038-1043 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[20]
郑德波, 杨小红, 李建生, 严建兵, 张士龙, 贺正华, 黄益勤. 基于SNP标记的玉米株高及穗位高QTL定位. 作物学报, 2013, 39: 549-556ZhengD B, YangX H, LiJ S, YanJ B, ZhangS L, HeZ H, HuangY Q. QTL identification for plant height and ear height based on SNP mapping in maize (Zea mays L. ). Acta Agron Sin, 2013, 39: 549-556 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[21]
苗晗, 顾兴芳, 张圣平, 张忠华, 黄三文, 王烨. 利用永久群体在不同环境下定位黄瓜株高QTL. 中国农业科学, 2012, 45: 4552-4560MiaoH, GuX F, ZhangS P, ZhangZ H, HuangS W, WangY. Detection of QTL for plant height in different environments using an RIL populution in cucumber. Sci Agric Sin, 2012, 45: 4552-4560 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[22]
张维峰, 赵守贤. 国外蓖麻育种与遗传研究动态. 中国油料作物作物学报, 1988, (1): 90-92ZhangW F, ZhaoS X. Foreign breeding and genetic research trends of castor. Chin J Oil Crop Sci, 1988, (1): 90-92 (in Chinese)[本文引用:1][CJCR: 0.95]
[23]
姚远, 李凤山, 陈永胜, 李金琴, 黄风兰, 王永佳. 国内外蓖麻研究动态. 内蒙古民族大学学报, 2009, 24(2): 172-174YaoY, LiF S, ChenY S, LiJ Q, HuangF L, WangY J. Research progress on castor. J Inner Mongolia Univ Nat, 2009, 24(2): 172-174 (in Chinese with English abstract)[本文引用:1]
[24]
GroosC, RobertN, BervasE, CharmetG. Genetic analysis of grain protein-content, grain yield and thousand -kernel weight in bread wheat. Theor Appl Genet, 2003, 106: 1032-1040[本文引用:1][JCR: 3.658]
[25]
张坤普, 徐宪斌, 田纪春. 小麦籽粒产量及穗部相关性状的QTL定位. 作物学报, 2009, 35: 270-278ZhangK P, XuX B, TianJ C. QTL mapping for grain yield and spike related traits in common wheat. Acta Agron Sin, 2009, 35: 270-278 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[26]
李文福, 刘宾, 彭涛, 袁倩倩, 韩淑晓, 田纪春. 利用DH和IF2两个群体进行小麦粒重、粒型和硬度的QTL分析. 中国农业科学, 2012, 45: 3453-3462LiW F, LiuB, PengT, YuanQ Q, HanS X, TianJ C. Detection of QTL for kernel weight, grain size, and grain hardness in wheat using DH and immortalized F2 population. Sci Agric Sin, 2012, 45: 3453-3462 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[27]
覃鸿妮, 晏萌, 王召辉, 郭莹, 王辉, 孙海燕, 刘志斋, 蔡一林. 玉米籽粒中花色苷和黑色素含量的QTL分析. 作物学报, 2012, 38: 275-284QinH N, YanM, WangZ H, GuoY, WangH, SunH Y, LiuZ Z, CaiY L. QTL mapping for anthocyanin and melanin contents in maize kernel. Acta Agron Sin, 2012, 38: 275-284 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[28]
范冬梅, 杨振, 马占洲, 曾庆力, 杜翔宇, 蒋洪蔚, 刘春燕, 韩冬伟, 栾怀海, 裴宇峰, 陈庆山, 胡国华. 多环境条件下大豆倒伏性相关形态性状的QTL分析. 中国农业科学, 2012, 45: 3029-3039FanD M, YangZ, MaZ Z, ZengQ L, DuX Y, JiangH W, LiuC Y, HanD W, LuanH H, PeiY F, ChenQ S, HuG H. QTL analysis of lodging-resistance related traits in soybean in different environments. Sci Agric Sin, 2012, 45: 3029-3039 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
 , 毕川, 文淡悠, 郑军, 刘帅, 石卓兴, 成粤湘
, 毕川, 文淡悠, 郑军, 刘帅, 石卓兴, 成粤湘

{kind=link}
{kind=link}