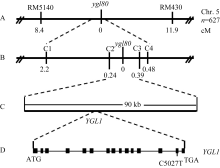
摘要通过化学诱变获得遗传稳定的水稻黄绿叶突变体ygl80。与野生型亲本10079相比,ygl80突变体在苗期和孕穗期叶片叶绿素分别下降76.64%和54.59%, 类胡萝卜素含量分别下降53.85%和41.18%, 成熟期株高、每株有效穗数、每穗着粒数、穗长和千粒重分别减少14.8%、16.5%、21.3%、9.1%和7.4%。遗传分析表明,ygl80的突变性状由1对隐性核基因控制。利用(ygl80/浙辐802) F2作为定位群体, 将突变基因定位在第5染色体长臂InDel标记C2和C3之间, 遗传距离分别为0.24 cM 和0.39 cM, 两标记之间的物理距离约为90 kb, 此区间内包含11个预测基因。基因组序列分析发现,ygl80突变体在编码叶绿素合酶的YGL1 (LOC_Os05g28200)基因编码区第5027碱基处(位于第14外显子), 碱基C突变为碱基T, 使编码蛋白序列第348位的脯氨酸(Pro)突变成亮氨酸(Leu)。该基因是已报道的水稻ygl1黄绿叶突变基因的等位基因。ygl80突变体在整个生育期都表现为黄绿叶, 而ygl1突变体在苗期叶片黄化, 中期慢慢转绿, 后期叶色以及总叶绿素和类胡萝卜素的含量接近野生型, 这可能是YGL1基因编码的叶绿素合酶蛋白的氨基酸不同突变位点造成的。
关键词:水稻; 黄绿叶突变体; YGL1; 遗传分析; 精细定位 Genetic Analysis and Gene Fine Mapping of Yellow-Green Leaf Mutantygl80 in Rice LI Yan-Qun1,**, GAO Jia-Xu1,**, XIAO Yun-Hua1, LI Xiu-Lan2, PU Xiang1, SUN Chang-Hui1, WANG Ping-Rong1, DENG Xiao-Jian1,* 1 Rice Research Institute, Sichuan Agricultural University, Chengdu 611130, China
2 College of Life Science, Qufu Normal University, Qufu 273165, China
Fund: AbstractA yellow-green leaf mutantygl80 was isolated by chemical mutagenesis. Compared with the wild-type parent 10079, chlorophyll content of theygl80mutant decreased by 76.64% and 54.59%, and the carotenoid content decreased by 53.85% and 41.18% at the seedling and booting stages, respectively. In addition, plant height, number of productive panicles per plant, number of spikelets per panicle, panicle length and 1000-grain weight reduced by 14.8%, 16.5%, 21.3%, 9.1%, and 7.4%, respectively, at the maturity. Genetic analysis showed that the yellow-green leaf trait of theygl80mutant was controlled by one pair of recessive nuclear genes. Genetic mapping of the mutant gene was conducted by using 627 yellow-green leaf individuals from the F2 mapping population ofygl80/Zhefu 802. Finally, the mutant gene was mapped between InDel markers C2 and C3 on the long arm of chromosome 5, with genetic distances of 0.24 cM and 0.39 cM, respectively, and with physical distance of 90 kb, in this region eleven predicted genes had been annotated. Sequencing analysis of these candidate genes between the mutant and its wild-type parent revealed a single base change (C5027T) ofYGL1(LOC_Os05g28200) gene for chlorophyll synthase resulted in a missense mutation (P348L) in the encoded product, suggesting that theygl80mutant gene is allelic to theygl1gene. Theygl80 mutant exhibited yellow-green trait throughout the growing period. But theygl1mutant showed yellow-green trait at seedling stage, then turned into green slowly, and its leaf color and chlorophyll and carotenoid contents almost closed to those of the wild-type parent during the later stage of growth. Different phenotypes of the two mutants may be caused by different mutational sites of genomic sequence ofYGL1 gene encoding chlorophyll synthase.
Keyword:Oryza sativa L.; Yellow-green leaf mutant; YGL1; Genetic analysis; Fine mapping Show Figures Show Figures
图1 突变体 ygl80(右)与其野生型亲本10079(左)在苗期(A)及拔节期(B)植株形态Fig. 1 Plant phenotype of ygl80(right) and its wild-type parent 10079 (left) at the seedling (A) and elongation (B) stages
表1 突变体 ygl80与其野生型亲本10079主要农艺性状的比较 Table 1 Comparison of major agronomic traits between the ygl80 mutant and its wild-type parent 10079
性状 Trait
10079 (CK)
ygl80
比对照增减 Compared to CK (%)
抽穗期Days to heading (d)
119.0±0.6
121.0±1.0
1.7
株高Plant height (cm)
101.9±0.5
86.8±1.6
-14.8*
每株有效穗数Number of productive panicles per plant
11.5±0.5
9.6±0.6
-16.5*
穗长Panicle length (cm)
19.7±0.5
17.9±0.2
-9.1*
每穗着粒数Number of spikelets per panicle
111.5±5.9
87.8±5.6
-21.3*
结实率Seed setting rate (%)
91.9±1.9
89.0±1.3
-2.9
千粒重1000-grain weight (g)
24.3±0.5
22.5±0.1
-7.4*
*在0.05水平上差异显著。*Significantly different at 0.05 probability level.
表1 突变体 ygl80与其野生型亲本10079主要农艺性状的比较 Table 1 Comparison of major agronomic traits between the ygl80 mutant and its wild-type parent 10079
表2 Table 2 表2(Table 2)
表2 苗期和孕穗期 ygl80突变体与其野生型亲本10079叶片光合色素含量的比较 Table 2 Comparison of photosynthetic pigment contents between the ygl80 mutant and its wild-type parent 10079 during seedling and booting stages
生育期 Growth stage
材料 Material
叶绿素 Chl (mg g-1)
叶绿素a Chl a (mg g-1)
叶绿素b Chl b (mg g-1)
叶绿素a/b Chl a/b
β-胡萝卜素 β-Car (mg g-1)
苗期 Seedling stage
10079 (CK)
4.11±0.02
3.22±0.02
0.89±0.00
3.62±0.02
0.91±0.01
ygl80
0.96±0.00
0.89±0.00
0.07±0.00
12.71±0.18
0.42±0.00
比对照增减Compared to CK
-76.64%**
-72.36%**
-92.13%**
9.09**
-53.85 %**
孕穗期 Booting stage
10079
2.07±0.04
1.61±0.05
0.46±0.01
3.50±0.16
0.51±0.03
ygl80
0.94±0.04
0.81±0.03
0.13±0.01
6.23±0.83
0.30±0.02
比对照增减Compared to CK
-54.59%**
-49.69%**
-71.74 %**
2.73**
-41.18%**
*在0.05水平上差异显著。*Significantly different at 0.05 probability level.
表2 苗期和孕穗期 ygl80突变体与其野生型亲本10079叶片光合色素含量的比较 Table 2 Comparison of photosynthetic pigment contents between the ygl80 mutant and its wild-type parent 10079 during seedling and booting stages
KrolM, SpangfortM D, Huner N P A, OquistG, GustafssonP, JanssonS. Chlorophyll a/b-binding proteins, pigment conversions, and early light-induced proteins in a chlorophyll b-less barley mutant. Plant Physiol, 1995, 107: 873-883[本文引用:1][JCR: 6.555]
[3]
FalbelT G, MeehlJ B, StaehelinL A. Severity of mutant phenotype in a series of chlorophyll-deficient wheat mutants depends on light intensity and the severity of the block in chlorophyll synthesis. Plant Physiol, 1996, 112: 821-832[本文引用:1][JCR: 6.555]
[4]
CarolP, StevensonD, BisanzC, BreitenbachJ, Sand mannG, MacheR, Coupland G, KuntzM. Mutations in the Arabidopsis gene IMMUTANTS cause a variegated phenotype by inactivating a chloroplast terminal oxidase associated with phytoene desaturation. Plant Cell, 1999, 11: 57-68[本文引用:1][JCR: 9.251]
[5]
胡忠, 彭丽萍, 蔡永华. 一个黄绿色的水稻细胞核突变体. 遗传学报, 1981, 8: 256-261HuZ, PengL P, CaiY H. A yellow-green nucleus mutant of rice. Acta Genet Sin, 1981, 8: 256-261 (in Chinese with English abstract)[本文引用:1]
[6]
ZhaoY, DuL F, YangS H, LiS C, ZhangY Z. Chloroplast composition and structure differences in a chlorophyll-reduced mutant of oilseed rape seedlings. Acta Bot Sin, 2001, 43: 877-880[本文引用:1][JCR: 0.599]
[7]
NagataN, TanakaR, SatohS, TanakaA. Identification of a vinyl reductase gene for chlorophyll synthesis in Arabidopsis thaliana and implications for the evolution of prochlorococcus species. Plant Cell, 2005, 17: 233-240[本文引用:1][JCR: 9.251]
[8]
BealeS I. Green genes gleaned. Trends Plant Sci, 2005, 10: 309-312[本文引用:1][JCR: 11.808]
[9]
JungK H, HurJ, RyuC H, ChoiY, ChungY Y, MiyaoA, HirochikaH, AnG. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol, 2003, 44: 463-472[本文引用:1][JCR: 4.134]
[10]
ZhangH T, LiJ J, YooJ H, YooS C, ChoS H, KohH J, SeoH S, PaekN C. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol, 2006, 62: 325-337[本文引用:1][JCR: 3.518]
[11]
WangP R, GaoJ X, WanC M, ZhangF T, XuZ J, HuangX Q, SunX Q, DengX J. Divinyl chlorophyll(ide) a can be converted to monovinyl chlorophyll(ide) a by a divinyl reductase in rice. Plant Physiol, 2010, 153: 994-1003[本文引用:1][JCR: 6.555]
[12]
SakurabaY, RahmanM L, ChoS H, KimY S, KohH J, YooS C, PeakN C. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J, 2013, 74: 122-133[本文引用:2][JCR: 6.582]
[13]
WuZ M, ZhangX, HeB, DiaoL P, ShengS L, WangJ L, GuoX P, SuN, WangL F, JiangL, WangC M, ZhaiH Q, WanJ M. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:6][JCR: 6.555]
[14]
LeeS, KimJ H, YooE S, LeeC H, HirochikaH, AnG. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol Biol, 2005, 57: 805-818[本文引用:1][JCR: 3.518]
[15]
LichtenthalerH K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Method Enzymol, 1987, 148: 350-382[本文引用:1][JCR: 2.002]
[16]
McCouchS R, KochertG, YuZ H, WangY Z, KhushG S, CoffmanW R, TanksleyS D. Molecular mapping of rice chromosomes. Theor Appl Genet, 1988, 76: 815-829[本文引用:1][JCR: 3.658]
[17]
PanaudO, ChenX, McCouchS R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L. ). Mol Gen Genet, 1996, 252: 597-607[本文引用:1]
[18]
TanakaR, TanakaA. Tetrapyrrole biosynthesis in higher plants. Annu Rev Plant Biol, 2007, 58: 321-346[本文引用:1][JCR: 25.962]
[19]
MarkwellJ P, ThornberJ P, BoggsR T. Higher plant chloroplasts: evidence that all the chlorophyll exists as chlorophyll-protein complexes. Proc Natl Acad Sci USA, 1979, 76: 1233-1235[本文引用:1][JCR: 9.737]
[20]
LiuW Z, FuY P, HuG C, SiH M, ZhuL, WuC, SunZ X. Identification and fine mapping of a thermo-sensitive chlorophyll deficient mutant in rice (Oryza sativa L. ). Planta, 2007, 226: 785-795[本文引用:1][JCR: 3.347]
[21]
EspinedaC E, Linford, AS, DevineD, BrusslanJ A. The AtCAO gene, encoding chlorophyll a oxygenase, is required for chlorophyll b synthesis in Arabidopsis thaliana. Proc Natl Acad Sci USA, 1999, 96: 10507-10511[本文引用:2][JCR: 9.737]
[22]
RüdigerW. Biosynthesis of chlorophyll b and the chlorophyll cycle. Photosynth Res, 2002, 74: 187-193[本文引用:1][JCR: 3.15]
[23]
OsterU, TanakaR, TanakaA, RüdigerW. Cloning and functional expression of the gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabidopsis thaliana. Plant J, 2000, 21: 305-310[本文引用:1][JCR: 6.582]
[24]
KusabaM, ItoH, MoritaR, IidaS, SatoY, FujimotoM, KawasakiS, TanakaR, HirochikaH, NishimuraM, TanakaA. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell, 2007, 19: 1362-1375[本文引用:1][JCR: 9.251]
[25]
MeguroM, ItoH, TakabayashiA, TanakaR, TanakaA. Identification of the 7-hydroxymethyl chlorophyll a reductase of the chlorophyll cycle in Arabidopsis. Plant Cell, 2011, 23: 3442-3453[本文引用:1][JCR: 9.251]
[26]
OsterU, BauerC E, RüdigerW. Characterization of chlorophyll a and bacteriochlorophyll a synthases by heterologous expression in Escherichia coli. J Biol Chem, 1997, 272: 9671-9676[本文引用:1][JCR: 4.651]
[27]
SollJ, SchultzG, RüdigerW, BenzJ. Hydrogenation of geranylgeraniol: two pathways exist in spinach chloroplasts. Plant Physiol, 1983, 71: 849-854[本文引用:1][JCR: 6.555]
[28]
SchmidH C, OsterU, KögelJ, LenzS, RüdigerW. Cloning and characterisation of chlorophyll synthase from Avena sativa. Biol Chem, 2001, 382: 903-911[本文引用:1][JCR: 2.683]
[29]
OsterU, RüdigerW. The G4 gene of Arabidopsis thaliana encodes a chlorophyll synthase of etiolated plants. Bot Acta, 1997, 110: 420-423[本文引用:1][JCR: 0.374]
[30]
吴自明, 张欣, 万建民. 水稻黄绿叶基因的克隆及应用. 生命科学, 2007, 19: 614-615WuZ M, ZhangX, WanJ M. Cloning and application of yellow-green leaf gene in rice. Chin Bull Life Sci, 2007, 19: 614-615 (in Chinese with English abstract)[本文引用:2][CJCR: 0.577]





{kind=link}
{kind=link}