关键词:白菜型冬油菜; Cu/Zn-SOD基因克隆; 低温; 表达分析; SOD活性; 低温诱导蛋白 Cloning and Expression Analysis of Copper and Zinc Superoxide Dismutase (Cu/Zn-SOD) Gene fromBrassica campestris L. ZENG Xiu-Cun1,2, LIU Zi-Gang2, SHI Peng-Hui2, XU Yao-Zhao1, SUN Jia3, FANG Yan4, YANG Gang2, WU Jun-Yan2, KONG De-Jing2, SUN Wan-Cang2,* 1College of Agronomy and Biotechnology, Hexi University, Zhangye 734000, China
2College of Agronomy, Gansu Agricultural University / Gansu Engineering Research Center of Rapeseed, Lanzhou 730070, China
3Department of Plant Sciences, University of Saskatchewan, Saskatoon, Saskatchewan S7N5A8, Canada
4Research and Testing Center of Gansu Agricultural University, Lanzhou 730070, China
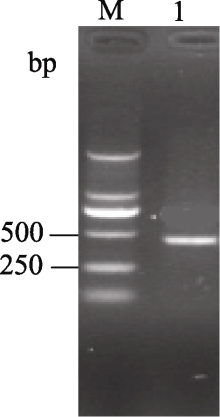
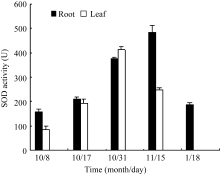
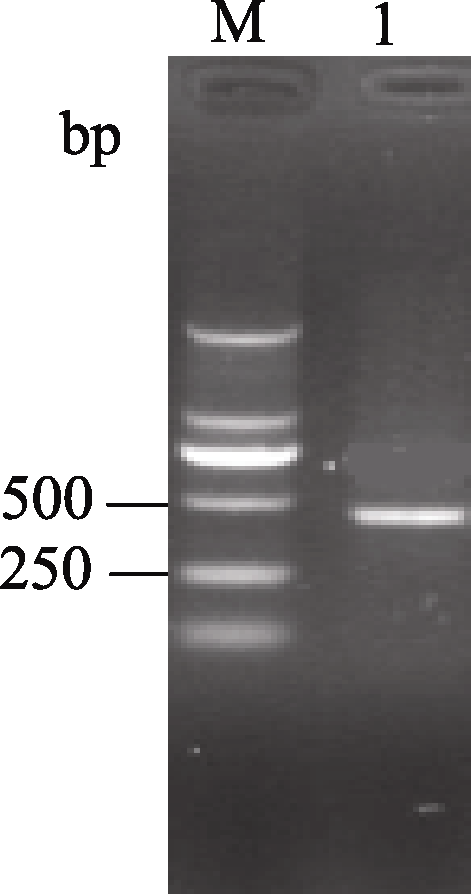
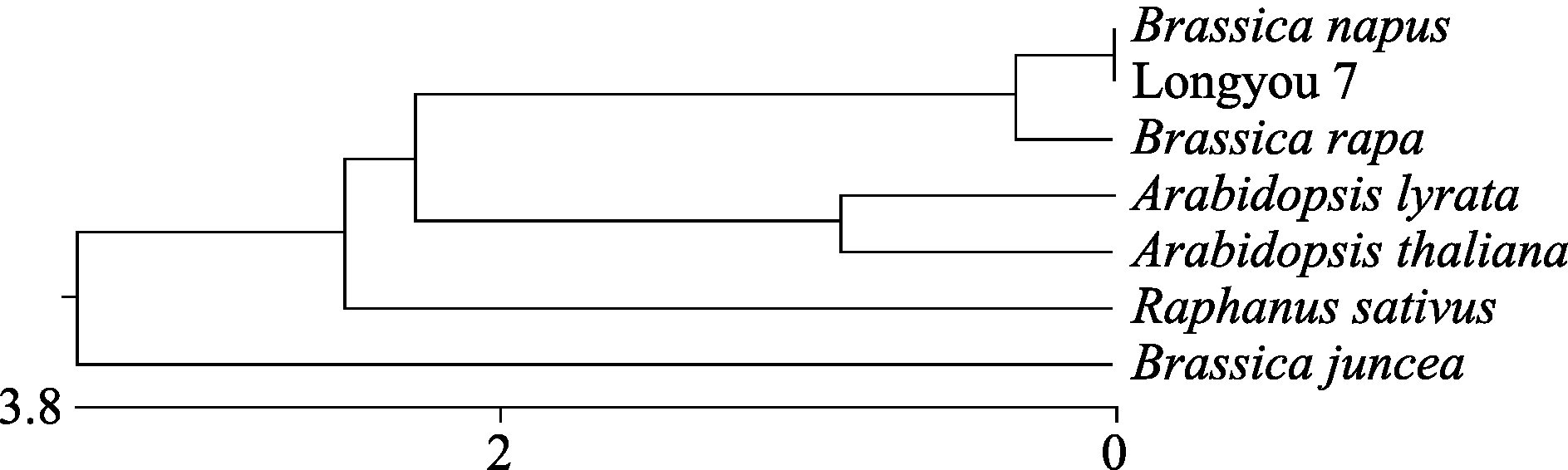
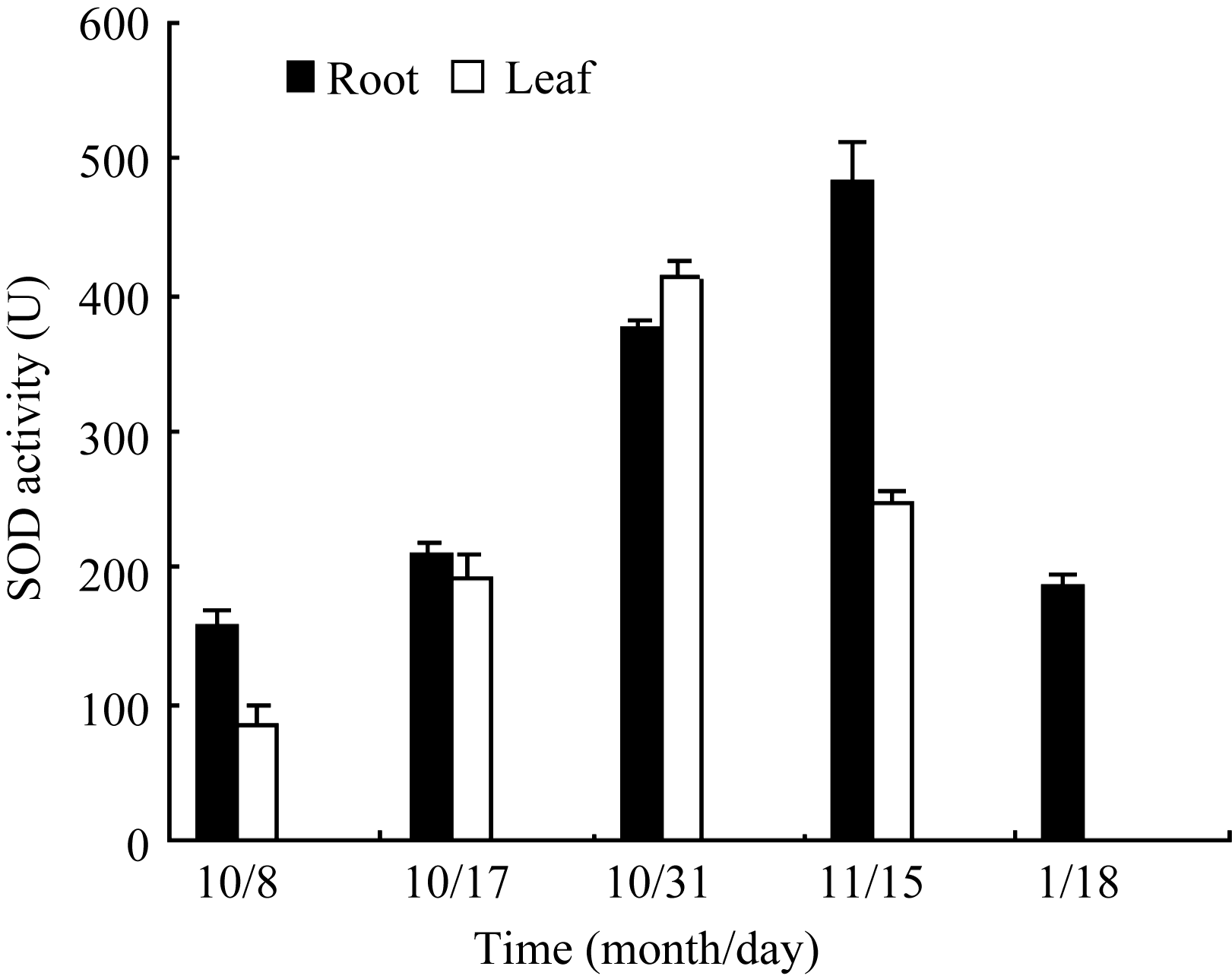
Fund: AbstractSuperoxide Dismutase (SOD) is a key enzyme eliminating reactive oxygen species (ROS), and the copper and zinc superoxide dismutase (Cu/Zn-SOD) is the most important active oxygen scavenger in SOD family, which can improve plant tolerance to environmental stresses. The cDNA ofBrassica campestris L. cultivar Longyou 7 was cloned by RT-PCR, using the primers designed according to the published cruciferCu/Zn-SOD cDNA sequences. The sequence ofCu/Zn-SODfromB. campestris L. was 459 bp, encoding a predicted protein of 152 amino acid residues. Bioinformatics analysis showed that amino sequence similarity withBrassica napus was 99%, and the predicted Cu/Zn-SOD protein was a hydrophilic protein without signal-peptide as well as transmembrane region. It contained specific sequence characteristics and conserved domain of copper and zinc superoxide dismutase (Cu/Zn-SOD) superfamily. The expression analysis ofCu/Zn-SOD gene using semi-quantitative RT-PCR and results of SOD activity in response to lower temperature showed that theCu/Zn-SODwas a differential expressed gene induced by lower temperature. Results of separating low-temperature-induced proteins fromBrassica campestris L. cultivar Longyou 6 by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and mass spectrometry further showed thatCu/Zn-SOD was a stress- responsed gene, whose expression was induced by lower temperature.
Keyword:Winter rape (Brassica compestris L.); Cu/Zn-SOD gene cloning; Lower temperature; Expression analysis; Activity of SOD; Low-temperature-induced protein Show Figures Show Figures
图2 Cu/Zn-SOD基因的编码区核酸序列及推导氨基酸序列方框为起始密码子ATG; *为终止密码子TAA。 ATG is indicated in box; TAA is marked by asterisk.Fig. 2 Nucleotide sequence of Cu/Zn-SODgene and its deduced amino acid sequence
图6 叶片低温诱导蛋白SDS-PAGE电泳图谱M: 标准蛋白; 1: 对照; 2~6: 依次为低温处理5、10、15、20和25 d蛋白; 7: 常温恢复生长10 d蛋白。Fig. 6 SDS-PAGE electrophoretogram of the low-temperature- induced protein in leafM: protein marker; 1: control; 2-6: SDS-PAGE of proteins from low temperature for 5, 10, 15, 20, and 25 days, respectively; 7: protein from room temperature for 10 days.
表1 Table 1 表1(Table 1)
表1 蛋白质串联质谱(LC-MS/MS)的数据库搜索鉴定结果 Table 1 Result of searching protein database using peptide sequenced from LC-MS/MS
蛋白质 条带 Protein
搜索条件Searching condition
搜索结果 Searching results
真实肽段 Ture peptide
物种 Species
匹配肽段 Match of fragment
离子分值 Score
蛋白质名称 Protein
物种 Species
序列号 No. of accession
得分 Score
D3
KGGHELSLTTGNAGGRL
Brassica campestris
KGGHELSLTTGNAGGRL
82
copper/zinc superoxide dismutase
Brassica rapa
gi|312837922
372
RAFVVHELKDDLGKG
RAFVVHELKDDLGKG
133
KGNSDVEGVVTLTQDDSGPTKV
KGNSDVEGVVTLTQDDSGPTKV
157
表1 蛋白质串联质谱(LC-MS/MS)的数据库搜索鉴定结果 Table 1 Result of searching protein database using peptide sequenced from LC-MS/MS
孙万仓, 马卫国, 雷建民, 刘秦, 杨仁义, 武军艳, 王学芳, 叶剑, 曾军, 张亚宏, 康艳丽, 郭秀娟, 魏文惠, 杨杰, 蒲媛媛, 曾潮武, 刘红霞. 冬油菜在西北旱寒区的适应性和北移的可行性研究. 中国农业科学, 2007, 40: 2716-2726SunW C, MaW G, LeiJ M, LiuQ, YangR Y, WuJ Y, WangX F, YeJ, ZengJ, ZhangY H, KangY L, GuoX J, WeiW H, YangJ, PuY Y, ZengC W, LiuH X. Study on adaptation and introduction possibility of winter rapeseed to dry and cold areas in northwest China. Sci Agric Sin, 2007, 40: 2716-2726 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[2]
姜海杨, 孙万仓, 曾秀存, 方彦, 陈姣荣, 史鹏辉, 赵彩霞, 何丽. 播期对北方白菜型冬油菜生长发育及产量的影响. 中国油料作物学报, 2012, 34: 620-626JiangH Y, SunW C, ZengX C, FangY, ChenJ R, ShiP H, ZhaoC X, HeL. Effect of sowing date on Brassica rapa growth and yield in northern China. Chin J Oil Crop Sci, 2012, 34: 620-626 (in Chinese with English abstract)[本文引用:1][CJCR: 0.95]
[3]
AlscherR G, ErturkN, HeathL S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plant. J Exp Bot, 2002, 53: 1331-1341[本文引用:1][JCR: 5.242]
[4]
AllenR D, WebbR P, SchakeS A. Use of transgenic plants to study antioxidant defenses. Free Radic Biol Med, 1997, 23: 473-479[本文引用:1][JCR: 5.271]
[5]
郭丽红, 吴晓岚, 龚明. 谷胱甘肽还原酶和超氧化物歧化酶在玉米幼苗热激诱导的交叉适应中的作用. 植物生理学通讯, 2005, 41: 429-432GuoL H, WuX L, GongM. Roles of glutathione reductase and superoxide dismutase in heat-shock-induced cross adaptation in maize seedlings. Plant Physiol Commun, 2005, 41: 429-432 (in Chinese with English abstract)[本文引用:1][CJCR: 0.849]
[6]
LawrenceV G, NicoleT B, WuG H, LuoX M, LiuX J, MichaelL G, AlanM H. Superoxide dismutase: an all-purpose gene for agri-biotechnology. Mol Breed, 2009, 24: 103-115[本文引用:1][JCR: 3.251]
[7]
AsadaK, KanematsuS, OkadaS, HayakanaT. Phylogenic distribution of three types of superoxide dismutase in organisms and cell organelles. In: Bannister J V, Hao H, eds. Chemical and Biochemical Aspects of Superoxide Dismutase, 1980. pp128-135[本文引用:1]
[8]
冯昌军, 罗新义, 沙伟, 王凤国. 低温胁迫对苜蓿品种幼苗SOD、POD活性和脯氨酸含量的影响. 草业科学, 2005, 22(6): 29-32FengC J, LuoX Y, ShaW, WangF G. Effect of low temperature stress on SOD, POD activity and proline content of alfalfa. Pratac Sci, 2005, 22(6): 29-32 (in Chinese with English abstract)[本文引用:1]
[9]
SongF N, YangC P, LiuX M, LiG B. Effect of salt stress on activity of superoxide dismutase (SOD) in Ulmus pumila L. J For Res, 2006, 17: 13-16[本文引用:1]
[10]
张怡, 罗晓芳, 沈应柏. 土壤逐渐干旱过程中刺槐新品种苗木抗氧化系统的动态变化. 浙江林学院学报, 2005, 22: 166-169ZhangY, LuoX F, ShenY B. Dynamic change s of anti-oxidation system in new cultvars of Robinia pseudoacacia under gradual drought stress of soil. J Zhejiang For Coll, 2005, 22: 166-169 (in Chinese with English abstract)[本文引用:1]
[11]
FaizeM, BurgosL, FaizeL, PiquerasA, NicolasE, Barba-Espin G, Clemente-MorenoM J, AlcobendasR, ArtlipT, Hernand ezJ A. Involvement of cytosolic ascorbate peroxidase and Cu/Zn-superoxide dismutase for improved tolerance against drought stress. J Exp Bot, 2011, 62: 2599-2613[本文引用:1][JCR: 5.242]
[12]
胡根海, 喻树迅, 范术丽, 宋美珍. 编码棉花胞质铜锌超氧物歧化酶基因的克隆与表达分析. 中国农业科学, 2007, 40: 1602-1609HuG H, YuS X, FanS L, SongM Z. Cloning and expressing of a gene encoding cytosolic copper/zinc superoxide dismutase in the upland cotton. Sci Agric Sin, 2007, 40: 1602-1609 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[13]
SumioK, KoziA. Characteristic of amino acid sequences of chloroplast and cytoplast isozymes of Cu/Zn superoxide dismutase in spinach, rice and horsetail. Plant Cell Physiol, 1990, 31: 99-112[本文引用:1][JCR: 4.134]
[14]
KernodleS P, Scand aliosJ G. A comparison of the structure and function of the highly homologous maize antioxidant Cu/Zn superoxide dismutase genes SOD4 and SOD4A. Genetics, 1996, 144: 317-328[本文引用:1][JCR: 4.389]
[15]
马旭俊, 朱大海. 植物超氧化物歧化(SOD)的研究进展. 遗传, 2003, 25: 225-231MaX J, ZhuD H. Functional roles of the plant superoxide dismutase. Hereditas, 2003, 25: 225-231 (in Chinese with English abstract)[本文引用:1][CJCR: 0.928]
[16]
MichelP, FatheyS. Synthesis of freezing tolerance proteins in leaves, crown, and roots during cold acclimation of wheat. Plant Physiol, 1989, 89: 577-585[本文引用:1][JCR: 6.555]
[17]
王瑞刚, 陈少良, 刘力源, 郝志勇, 翁海娇, 李鹤, 杨爽, 段杉. 盐胁迫下3种杨树的抗氧化能力与耐盐性研究. 北京林业大学学报, 2005, 27(3): 46-52WangR G, ChenS L, LiuL Y, HaoZ Y, WengH J, LiH, YangS, DuanS. Genotypic differences in antioxidative ability and salt tolerance of three poplars under salt stress. J Beijing For Univ, 2005, 27(3): 46-52 (in Chinese with English abstract)[本文引用:1]
[18]
HonW C, GriffithM, ChongP, Yang D S C. Extraction and isolation of antifreeze proteins from winter rye (Secale cereale L. ) leaves. Plant Physiol, 1994, 104: 971-980[本文引用:1][JCR: 6.555]
[19]
章慧慧, 励勤荣. 超氧化物歧化酶的研究和应用现状. 农产品加工(学刊), 2007, (8): 28-32ZhangH H, LiQ R. Study on Superoxide Dismutase. Acad Period Farm Prod Proc, 2007, (8): 28-32 (in Chinese with English abstract)[本文引用:1]
[20]
TainerJ A, GetzoffE D, BeemK M, RichardsonJ. Determination and analysis of the 2A structure of copper, zinc supero- xide dismutase. J Mol Biol, 1982, 160: 181-184[本文引用:1][JCR: 3.905]
[21]
纪砚耘, 化文平, 王喆之. 丹参铜锌超氧化物歧化酶(Cu/Zn-SOD)基因的克隆与生物信息学分析. 陕西师范大学学报(自然科学版), 2011, 39(3): 56-61JiY Y, HuaW P, WangZ Z. Cloning and bioinformatics analysis of Cu/Zn-SOD gene from Salvia miltiorrhiza Bunge. J Shaanxi Norm Univ (Nat Sci Edn), 2011, 39(3): 56-60 (in Chinese with English abstract)[本文引用:1]
[22]
KeS S. Effects of copper on the photosynthesis and oxidative metabolism of amaranthus tricolor seedlings. Agric Sci China, 2007, 6: 1182-1192[本文引用:1][JCR: 0.527][CJCR: 0.4431]
[23]
McKersieB D, MurnaghanJ, JonesK S, BowleyS R. Iron- superoxide dismutase expression in transgenic alfalfa increases winter survival without a detectable increase in photosynthetic oxidative stress tolerance. Plant Physiol, 2000, 122: 1427-1438[本文引用:1][JCR: 6.555]
[24]
MullineauxP M, CreissenG P. Glutathione reductase: regulation and role in oxidative stress. In: Scand alios J G, ed. Oxidative Stress and the Molecular Biology of Antioxidant Defenses. New York: Cold Spring Harbor Laboratory Press, 1997. pp667-713[本文引用:1]
[25]
李玉坤, 王学敏, 高洪文, 仁爱琴, 王赞, 孙桂枝. 东方山羊豆Cu/Zn-SOD基因的克隆及表达分析. 遗传, 2012, 34: 95-101LiY K, WangX M, GaoH W, RenA Q, WangZ, SunG Z. Cloning and expression analysis of Cu/Zn-SOD gene from Galega orientalis L. Hereditas (Beijing), 2012, 34: 95-101 (in Chinese with English abstract)[本文引用:1]
[26]
MallickN, MohnF H. Reactive oxygen species: response of algal cells. J Plant Physiol, 2000, 157: 183-193[本文引用:1][JCR: 2.699]
[27]
KnrzerO C, DurnerJ, BgerP. Alterations in the antioxidative system of suspension-cultured soybean cells (Glycine max) induced by oxidative stress. Physiol Plant, 1996, 97: 388-396[本文引用:1][JCR: 6.555]
[28]
XuJ, DuanX G, YangJ, JohnR B, ZhangP. Enhanced reactive oxygen species scavenging by overproduction of superoxide dismutase and catalase delays postharvest physiological deterioration of cassava storage roots. Plant Physiol, 2013, 161: 1517-1528[本文引用:1][JCR: 6.555]
[29]
HuL X, HuangZ H, LiuS Q, FuJ M. Growth response and gene expression in antioxidant-related enzymes in two bermudagrass genotypes differing in salt tolerance. J Am Soc Hort Sci, 2012, 137: 134-143[本文引用:1]
[30]
KacaokaT, DedaK. Heat-elable COR (cold-hegulated) proteins associated with freezing tolerance in epinacb. Plant Cell Physiol, 1992, 33: 1107-1114[本文引用:1][JCR: 4.134]
[31]
李生泉, 李锐, 范月仙. 棉苗抗冷性与低温诱导蛋白关系的研究. 棉花学报, 2006, 18: 43-46LiS Q, LiR, FanY X. Study on the relation between chilling resistance and low-temperature-induced proteins in cotton seeding. Cotton Sci, 2006, 18: 43-46 (in Chinese with English abstract)[本文引用:1][CJCR: 1.462]
[32]
HerouartD, VanMontagu M, InzéD. Redox-activated expression of the cytosolic copper/zinc superoxide dismutase gene in Nicotiana. Proc Natl Acad Sci USA, 1993, 90: 3108-3112[本文引用:1][JCR: 9.737]
[33]
WuG, WilenR W, RobertsonA J, GustaL V. Isolation, chromo-somal localization, and differential expression of mitochondrial manganese superoxide dismutase and chloroplastic copper/zinc superoxide dismutase genes in wheat. Plant Physiol, 1999, 120: 513-520[本文引用:1][JCR: 6.555]
[34]
杜秀敏, 殷文漩, 张慧, 赵彦修. 超氧化物歧化酶(SOD)研究进展. 中国生物工程杂志, 2003, 2(1): 48-50DuX M, YinW X, ZhangH, ZhaoY X. The researching progress of superoxide dismutase. J Chin Biotechnol, 2003, 2(1): 48-50 (in Chinese with English abstract)[本文引用:1][CJCR: 0.758]
[35]
AlscherR G, DonahueJ L, CramerC L. Reactive oxygen species and antioxidants relationships in green cells. Physiol Plant, 1997, 100: 224-233[本文引用:1][JCR: 6.555]







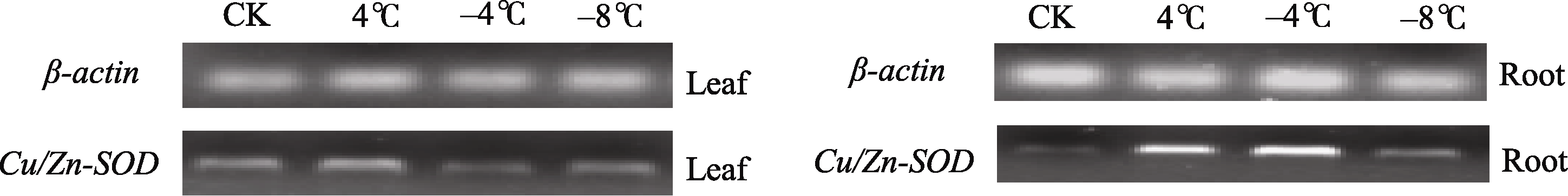


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}