关键词:宁麦9号; 宁麦13; SSR标记; 聚类分析; 遗传贡献 Genetic Contribution of Wheat Variety Ningmai 9 to Its Derivates JIANG Peng, CHEN Xiao-Lin, ZHANG Ping-Ping, ZHANG Peng, YAO Jin-Bao, MA Hong-Xiang* Jiangsu Provincial Key Laboratory of Agrobiology / Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China Fund: AbstractNingmai 9 is a soft wheat variety with high yield, wide adaption, and resistance to multiple diseases includingFusarium head blight, Yellow mosaic virus, powdery mildew, and sharp eye spots, which serves as a backbone parent in the southern Huaihe River valley. This study aimed to disclose the genetic contribution of Ningmai 9 to its derivates using SSR markers. A total of 471 alleles were detected on 170 SSR loci, with 1-6 alleles per locus and an average of 2.8. The UPGMA cluster analysis showed that Yangmai 18 and Ningmai 9 were clustered together firstly with the smallest genetic disrtance, and Yangfumai 4 and Ningmai 9 were clustered together with the largest genetic distance. Genetic similarity coefficient showed that more than a half of loci transferred from Ningmai 9 to its derived varieties. The percentage of alleles shared between Ningmai 9 and its derived varieties differed greatly among 21 chromosomes, but the average percentage was similar among A, B, and D genomes. Ten markers had the same amplification banding pattern in Ningmai 9 and nine derived varieties, and nine of them were linked to the QTLs or genes associated with important agronomic traits as previous reports. In comparison of genetic background between Ningmai 9 and Ningmai 13 (a direct selection line from Ningmai 9), the genetic similarity coefficient was 0.732, suggesting that the genetic back ground of Ningmai 13 is similar to that of Ningmai 9. Genotyping data showed that the markers associated with protein content and resistance to Fusarium head blight in Ningmai 13 were in correspondence with those in Ningmai 9. These results may partially explain the reason for elite agronomic traits of Ningmai 13 similar to Ningmai 9, such as resistance toFusarium head blight and soft wheat quality.
Keyword:Ningmai 9; Ningmai 13; SSR; Clustering analysis; Genetic contribution Show Figures Show Figures
图2 宁麦9号及其衍生品种的遗传连锁图灰色区域为宁麦13和宁麦9号相同的基因组区段, ◆表示宁麦9号与其衍生品种的相同标记位点。Fig. 2 Genetic linkage map of Ningmai 9 and its derivatesGray regions are identical linkage blocks between Ningmai 13 and Ningmai 9. ◆ shows the identical loci between Ningbai 9 and its derivates.
姚金保, 马鸿翔, 张平平, 姚国才, 杨学明, 任丽娟, 张鹏, 周淼平. 小麦优良亲本宁麦9号的研究与利用. 核农学报, 2012, 26: 17-21YaoJ B, MaH X, ZhangP P, YaoG C, YangX M, RenL J, ZhangP, ZhouM P. Research of wheat elite parent Ningmai 9 and its utilization. Acta Agric Nucl Sin, 2012, 26: 17-21 (in Chinese with English abstract)[本文引用:2]
[2]
YouG X, ZhangX Y, WangL F. An estimation of the minimum number of SSR loci needed to reveal genetic relationships in wheat varieties: Information from 96 rand om accessions with maximized genetic diversity. Mol Breed, 2005, 14: 397-406[本文引用:1][JCR: 3.251]
[3]
王兰芬, BalfourierF, 郝晨阳, ExbrayatF, 董玉琛, 盖红梅, 张学勇. 欧洲与东亚小麦品种遗传多样性的比较分析. 中国农业科学, 2007, 40: 2667-2678WangL F, BalfourierF, HaoC Y, ExbrayatF, DongY C, GeH M, ZhangX Y. Comparison of genetic diversity level between European and East-Asian wheat collections using SSR markers. Sci Agric Sin, 2007, 40: 2667-2678 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[4]
李玮瑜, 张斌, 张嘉楠, 昌小平, 李润植, 景蕊莲. 利用关联分析发掘小麦自然群体旗叶叶绿素含量的优异等位变异. 作物学报, 2012, 38: 962-970LiW Y, ZhangB, ZhangJ N, ChangX P, LiR Z, JingR L. Exploring elite alleles for chlorophyll content of flag leaf in natural population of wheat by association analysis. Acta Agron Sin, 2012, 38: 962-970 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[5]
HaoC Y, WangL F, GeH M, DongY C, ZhangX Y. Genetic diversity and linkage disequilibrium in Chinese bread wheat (Triticum aestivum L. ) revealed by SSR markers. PloS ONE, 2011, e6(2): e17279. DOI: 101371/journal. pone. 0017279[本文引用:1][JCR: 3.73]
[6]
HaoC Y, DongY C, WangL F, YouG X, ZhangH N, GeH M, JiaJ Z, ZhangX Y. Genetic diversity and construction of core collection in Chinese wheat genetic resources. Chin Sci Bull, 2008, 53: 1518-1526[本文引用:1][CJCR: 0.95]
[7]
孙慧敏, 张军, 赵团结, 盖钧镒. 亚洲地区中、外大豆品种幼苗期耐淹性与SSR标记的关联分析. 作物学报, 2010, 36: 1615-1623SunH M, ZhangJ, ZhaoT J, GaiJ Y. Association analysis between submergence tolerance and SSR markers in domestic and foreign soybean cultivars in Asia. Acta Agron Sin, 2010, 36: 1615-1623 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[8]
韩俊, 张连松, 李静婷, 石丽娟, 解超杰, 尤明山, 杨作民, 刘广田, 孙其信, 刘志勇. 小麦骨干亲本“胜利麦/燕大1817”杂交组合后代衍生品种遗传构成解析. 作物学报, 2009, 35: 1395-1404HanJ, ZhangL S, LiJ T, ShiL J, XieC J, YouM S, YangZ M, LiuG T, SunQ X, LiuZ Y. Molecular dissection of core parental cross “Triumph/Yand a 1817” and its derivatives in wheat breeding program. Acta Agron Sin, 2009, 35: 1395-1404 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[9]
盖红梅, 李玉刚, 王瑞英, 李振清, 王圣健, 高峻岭, 张学勇. 鲁麦14对山东新选育小麦品种的遗传贡献. 作物学报, 2012, 38: 954-961GeH M, LiY G, WangR Y, LiZ Q, WangS J, GaoJ L, ZhangX Y. Genetic contribution of Lumai 14 to novel wheat varieties developed in Shand ong Province. Acta Agron Sin, 2012, 38: 954-961 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[10]
徐鑫, 李小军, 李秀全, 杨欣明, 刘伟华, 高爱农, 李立会. 小麦骨干亲本"洛夫林10号"1BL/1RS在衍生品种中的遗传分析. 麦类作物学报, 2010, 30: 221-226XuX, LiX J, LiX Q, YangX M, LiuW H, GaoA N, LiL H. Inheritance of 1BL/1RS of founder parent Lovrin 10 in its progeny. J Triticeae Crops, 2010, 30: 221-226 (in Chinese with English abstract)[本文引用:1]
[11]
刘新伦, 司清林, 李琴琴, 王长有, 王亚娟, 张宏, 吉万全. 利用SSR标记分析小麦骨干亲本阿夫及衍生品种(系)的遗传多样性和变化趋势. 农业生物技术学报, 2012, 20: 983-995LiuX L, SiQ L, LiQ Q, WangC Y, WangY J, ZhangH, JiW Q. SSR analysis of genetic diversity and temporal trends of the core wheat (Triticum aestivum L. ) parent Funo and its derivative varieties (lines). J Agric Biotech, 2012, 20: 983-995 (in Chinese with English abstract)[本文引用:1][CJCR: 0.635]
[12]
袁园园, 王庆专, 崔法, 张景涛, 杜斌, 王洪刚. 小麦骨干亲本碧蚂4号的基因组特异位点及其在衍生后代中的传递. 作物学报, 2010, 36: 9-16YuanY Y, WangQ Z, CuiF, ZhangJ T, DuB, WangH G. Specific loci in genome of wheat milestone parent Bima 4 and their transmission in derivatives. Acta Agron Sin, 2010, 36: 9-16 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
SomersD J, IsaacP, EdwardsK. A high-density wheat microsatellite consensus map for bread wheat (Triticum aestivum L. ). Theor Appl Genet, 2004, 109: 1105-1114[本文引用:1][JCR: 3.658]
[15]
庄巧生. 中国小麦品种改良及其系谱分析. 北京: 中国农业出版社, 2003. pp10-13ZhuangQ S. Chinese Wheat Improvement and Pedigree Analysis. Beijing: China Agriculture Press, 2003. pp10-13(in Chinese)[本文引用:1]
[16]
姚金保, 姚国才, 杨学明, 马鸿翔, 张平平. 小麦收获指数遗传及其与农艺性状的相关分析. 江苏农业学报, 2008, 24: 5-10YaoJ B, YaoG C, YangX M, MaH X, ZhangP P. Inheritance of wheat harvest index and its correlations with agronomic traits. Jiangsu J Agric Sci, 2008, 24: 5-10 (in Chinese with English abstract)[本文引用:1][CJCR: 0.895]
[17]
姚金保, 任丽娟, 张平平, 杨学明, 马鸿翔, 姚国才, 张鹏, 周淼平. 小麦产量构成因素的双列杂交分析. 核农学报, 2011, 25: 633-638YaoJ B, RenL J, ZhangP P, YangX M, MaH X, YaoG C, ZhangP, ZhouM P. Diallel analysis for yield components of wheat. Acta Agric Nucl Sin, 2011, 25: 633-638 (in Chinese with English abstract)[本文引用:1]
[18]
姚金保, 张平平, 任丽娟, 杨学明, 马鸿翔, 姚国才, 张鹏, 周淼平. 软质冬小麦品种籽粒蛋白质含量的遗传分析. 江苏农业学报, 2011, 27: 469-474YaoJ B, ZhangP P, RenL J, YangX M, MaH X, YaoG C, ZhangP, ZhouM P. Inheritance of grain protein content in soft red wheat cultivars. Jiangsu J Agric Sci, 2011, 27: 469-474 (in Chinese with English abstract)[本文引用:1][CJCR: 0.895]
[19]
张平平, 姚金保, 马鸿翔. 小麦溶剂保持力的遗传分析. 江苏农业学报, 2010, 26: 1170-1175ZhangP P, YaoJ B, MaH X. Genetic analysis of solvent retention capacity in wheat. Jiangsu J Agric Sci, 2010, 26: 1170-1175 (in Chinese with English abstract)[本文引用:1][CJCR: 0.895]
[20]
姚金保, 任丽娟, 张平平, 杨学明, 马鸿翔, 姚国才, 张鹏, 周淼平. 小麦赤霉病的抗性遗传分析. 麦类作物学报, 2011, 31: 370-375YaoJ B, RenL J, ZhangP P, YangX M, MaH X, YaoG C, ZhangP, ZhouM P. Genetic analysis of resistance to Fusarium head blight in wheat. J Triticeae Crops, 2011, 31: 370-375 (in Chinese with English abstract)[本文引用:2]
[21]
周延清. DNA分子标记技术在植物研究中的应用. 北京: 化学工业出版社, 2005. pp174-178ZhouY Q. Application of DNA Markers in Plant Research. Beijing: Chemical Industry Press, 2005. pp174-178(in Chinese)[本文引用:1]
[22]
LiuS, PumphreyM O, GillB S, TrickH N, ZhangJ X, DolezeJ, ChalhoubB, AndersonJ A. Toward positional cloning of Fhb1, a major QTL for Fusarium head blight resistance in wheat. Cereal Res Commun, 2008, 36(suppl-B): 195-201[本文引用:1][JCR: 0.549]
[23]
张平平, 马庆, 钱存鸣, 马鸿翔. 宁麦13选系的遗传多样性及品质差异. 作物学报, 2008, 34: 1484-1488ZhangP P, MaQ, QianC M, MaH X. Genetic diversity and quality difference of systemic selective lines in Ningmai 13. Acta Agron Sin, 2008, 34: 1484-1488 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[24]
徐冠仁. 核农学导论. 北京: 原子能出版社, 1997. pp 104-10Xu G R. Introduction of Nuclear Agricultural Science. Atomic Energy Press, 1997. pp104-106(in Chinese)[本文引用:1]
[25]
RoyJ K, Band opadhyayR, RustgiS, BalyanH S, GuptaP K. Association analysis of agronomically important traits using SSR, SAMPL and AFLP markers in bread wheat. Curr Sci India, 2006, 90: 683-689[本文引用:1][JCR: 0.905]
[26]
YaoJ, WangL X, LiuL H, ZhaoC P, ZhengY L. Association mapping of agronomic traits on chromosome 2A of wheat. Genetica, 2009, 137: 67-75[本文引用:1]
[27]
周晓果, 景蕊莲, 郝转芳, 昌小平, 张正斌. 小麦幼苗根系性状的QTL分析. 中国农业科学, 2005, 38: 1951-1957ZhouX G, JingR L, HaoZ F, ChangX P, ZhangZ B. Mapping QTL for seedling root traits in common wheat. Sci Agric Sin, 2005, 38: 1951-1957 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[28]
EllisM H, RebetzkeG J, AzanzaF, RichardsR A, SpielmeyerW. Molecular mapping of gibberellin-responsive dwarfing genes in bread wheat. Theor Appl Genet, 2005, 111: 423-430[本文引用:1][JCR: 3.658]
[29]
LiY, SongY, ZhouR, BranlardG, JiaJ. Detection of QTLs for bread-making quality in wheat using a recombinant inbred line population. Plant Breed, 2009, 128: 235-243[本文引用:1][JCR: 1.175]
[30]
AnnapurnalillyB, BalgoundaH, MohinderP, SubhashB, SuryaprakasaR, ShubhadaT. Validation and identification of molecular markers linked to the leaf rust resistance gene Lr28 in wheat. J Appl Genet, 2011, 52: 171-175[本文引用:1][JCR: 1.847]
[31]
YangZ, GilbertJ, FedakG, SomersD J. Genetic characterization of QTL associated with resistance to Fusarium head blight in a doubled-haploid spring wheat population. Genome, 2005, 48: 187-196[本文引用:1][JCR: 1.668]
[32]
景蕊莲, 昌小平, 贾继增. 一个小麦不孕小穗数QTL的染色体7A的作图区间Xgwm260-WMC83-WMC301. 中国专利, CN200310115364. 6. 2005-05-25JingR L, ChangX P, JiaJ Z. Mapping interval Xgwm260- WMC83-WMC301 harboring a QTL for sterile spikelet number on chromosome 7A of wheat: China’s Patent No. CN2003 10115364. 6. 2005-05-25 (in Chinese)[本文引用:1]
[33]
ClarkeF R, KnoxR E, ClarkeJ M, PozniakC J. Quantitative trait loci for agronomic, pest resistance and end-use quality traits in a durum wheat doubled haploid population. In: Molina-Cano J L, Christou P, Graner A, Hammer K, Jouve N, Keller B, Lasa J M, Powell W, Royo C, Shewry P, Stanca A M, eds. Cereal Science and Technology for Feeding Ten Billion People: Genomics Era and Beyond. Zaragoza: CIHEAM/IRTA, 2008. pp111-113[本文引用:1]
[34]
JaiswalV, MirR R, MohanA, BalyanH S, GuptaP K. Association mapping for pre-harvest sprouting tolerance in common wheat (Triticum aestivum L. ). Euphytica, 2012, 188: 89-102[本文引用:1][JCR: 1.643]
[35]
LinF, XueS L, ZhangZ Z, ZhangC Q, KongZ X, YaoG Q, TianD G, ZhuH L, LiC J, CaoY, WeiJ B, LuoQ Y, MaZ Q. Mapping QTL associated with resistance to Fusarium head blight in the Nand a 2419×Wangshuibai population: II. Type I resistance. Theor Appl Genet, 2006, 112: 528-535[本文引用:1][JCR: 3.658]
[36]
GroosC, RobertN, BervasE, CharmetG. Genetic analysis of grain protein content, grain yield and thousand -kernel weight in bread wheat. Theor Appl Genet, 2003, 106: 1032-1040[本文引用:1][JCR: 3.658]
[37]
PrasadM, KumarN, KulwalP, RöderM, BalyanH, DhaliwalH, GuptaP. QTL analysis for grain protein content using SSR markers and validation studies using NILs in bread wheat. Theor Appl Genet, 2003, 106: 659-667[本文引用:1][JCR: 3.658]
 , 陈小霖, 张平平, 张鹏, 姚金保, 马鸿翔
, 陈小霖, 张平平, 张鹏, 姚金保, 马鸿翔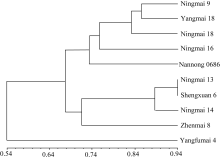



{kind=link}
{kind=link}