关键词:花生; 果糖-1; 6-二磷酸醛缩酶; 克隆; 系统发育分析; 非生物胁迫; 荧光定量PCR Cloning and Expression of Fructose-1,6-Bisphosphate Aldolase GeneAhFBA1 in Peanut (Arachis hypogaeaL.) CHEN Na1, PAN Li-Juan1, CHI Xiao-Yuan1,2, CHEN Ming-Na1, WANG Tong1, WANG Mian1, YANG Zhen1, HU Dong-Qing3, WANG Dao-Yuan4, YU Shan-Lin1,* 1Shandong Peanut Research Institute, Qingdao 266100, China
2Key Laboratory of Biology and Genetic Improvement of Oil Crops, Ministry of Agriculture, Oil Crops Research Institute, Chinese Academy of Agricultural Sciences, Wuhan, 430062, China
3Qingdao Entry-Exit Inspection and Quarantine Bureau, Qingdao 266001, China
4Zhangqiu Agriculture Bureau, Jinan 250200, China
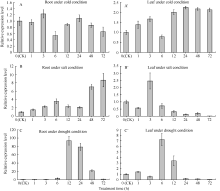
Fund: AbstractIn this article, a fructose-1,6-bisphosphate aldolase (FBA) gene was cloned from the leaf of peanut (Arachis hypogaea L. cultivar Huayu 33) using RT-PCR, and was designated asAhFBA1. The whole sequence ofAhFBA1 is 1489 bp and its open reading frame is 1200 bp, encoding a polypeptide of 400 amino acids. Its protein was predicted to be located in chloroplast, containing the conserved glycolytic domain. Multiple sequence alignments and phylogenetic analysis of FBA proteins indicated AhFBA1 was most similar with FBA fromGlycine max, Medicago truncatula,Cicer arietinum, andPhaseolus vulgaris. The results of Real-time RT-PCR showed that the expression ofAhFBA1 was induced distinctly in both peanut root and leaf under salt and drought conditions, suggesting thatAhFBA1 may participate in the salt and drought stress regulation of peanut. The expression ofAhFBA1 was also induced by exogenous ABA in both peanut leaf and root, which indicated thatAhFBA1 may regulate peanut abiotic stresses resistance through ABA-dependent pathway.
Keyword:Peanut; Fructose-1; 6-bisphosphate aldolase; Clone; Phylogenetic analysis; Abiotic stresses; Real-time PCR Show Figures Show Figures
JangJ C, LeonP, ZhouL, SheenJ. Hexokinase as a sugar sensor in higher plants. Plant Cell, 1997, 9: 5-19[Cited within: 1][JCR: 9.251]
[2]
LoretiE, BellisL, AlpiA, PerataP. Why and how do plant cells sense sugars?Ann Bot, 2001, 88: 803-812[Cited within: 1][JCR: 0.657]
[3]
FolgadoR, SergeantK, RenautJ, SwennenR, HausmanJ F, PanisB. Changes in sugar content and proteome of potato in response to cold and dehydration stress and their implications for cryopreservation. J Proteomics, 2014, 98: 99-111[Cited within: 1][JCR: 4.088]
ChoY H, YooS D. Signaling role of fructose mediated by FINS1/FBP in Arabidopsis thaliana. PLoS Genet, 2011, 7: e1001263[Cited within: 1][JCR: 8.517]
[6]
YamadaS, KomoriT, HashimotoA, KuwataS, ImasekiH, KuboT. Differential expression of plastidic aldolase genes in Nicotiana plants under salt stress. Plant Sci, 2000, 154: 61-69[Cited within: 3][JCR: 2.922]
[7]
JiangY, YangB, HarrisN S, DeyholosM K. Comparative proteomic analysis of NaCl stress-responsive proteins in Arabidopsis roots. J Exp Bot, 2007, 58: 3591-3607[Cited within: 1][JCR: 5.242]
[8]
NdimbaB K, ChivasaS, SimonW J, SlabasA R. Identification of Arabidopsis salt and osmotic stress responsive proteins using two-dimensional difference gel electrophoresis and mass spectrometry. Proteomics, 2005, 5: 4185-4196[Cited within: 1][JCR: 4.132]
[9]
ProvartN J, GilP, ChenW, HanB, ChangH S, WangX, ZhuT. Gene expressionphenotypes of Arabidopsis associated with sensitivity to low temperatures. Plant Physiol, 2003, 132: 893-906[Cited within: 1][JCR: 6.555]
[10]
LuW, TangX, HuoY, XuR, QiS, HuangJ, ZhengC, WuC A. Identification and characterization of fructose 1, 6-bisphosphate aldolase genes in Arabidopsis reveal a gene family with diverse responses to abiotic stresses. Gene, 2012, 503: 65-74[Cited within: 1][JCR: 2.196]
[11]
FanW, ZhangZ L, ZhangY L. Cloning and molecular characterization of fructose-1, 6-bisphosphate aldolase gene regulated by high-salinity and drought in Sesuvium portulacastrum. Plant Cell Rep, 2009, 28: 975-984[Cited within: 2][JCR: 2.509]
[12]
ChenM, MishraS, HeckathornS A, FrantzJ M, KrauseC. Proteomic analysis of Arabidopsis thaliana leaves in response to acute boron deficiency and toxicity reveals effects on photosynthesis, carbohydrate metabolism, and protein synthesis. J Plant Physiol, 2014, 171: 235-242[Cited within: 1][JCR: 2.699]
[13]
KochK. Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr Opin Plant Biol, 2004, 7: 235-246[Cited within: 1][JCR: 8.455]
[14]
HoekstraF A, CroweL M, CroweJ H. Differential desiccation sensitivity of corn and pennisetum pollen linked to their sucrose contents. Plant Cell Environ, 1989, 12: 83-91[Cited within: 1][JCR: 5.135]
[15]
SmeekensS, RookF. Sugar sensing and sugar-mediated signal transduction in plants. Plant Physiol, 1997, 115: 7-13[Cited within: 1][JCR: 6.555]
[16]
Rolland F, Baena-GonzalezE, SheenJ. Sugar sensing and signaling in plants: conserved and novel mechanisms. Annu Rev Plant Biol, 2006, 57: 675-709[Cited within: 1][JCR: 25.962]
[17]
HansonH D, HitzW D. Metabolic responses of mesophytes to plant water deficits. Annu Rev Plant Physiol, 1982, 33: 163-203[Cited within: 1]
[18]
WanX, MoA, LiuS, YangL, LiL. Constitutive expression of a peanut ubiquitin-conjugating enzyme gene in Arabidopsis confers improved water-stress tolerance through regulation of stress- responsive gene expression. J Biosci Bioeng, 2011, 111: 478-484[Cited within: 1][JCR: 1.737]
[19]
WangT, ChenX, ZhuF, LiH, LiL, YangQ, ChiX, YuS, LiangX. Characterization of peanut germin-like proteins, AhGLPs in plant development and defense. PLoS One, 2013, 8: e61722[Cited within: 1][JCR: 3.73]
[20]
万书波. 中国花生栽培学. 上海: 上海科学技术出版社, 2003. pp16-20WanS B. Peanut Cultivation in China. Shanghai: Shanghai Scientific and Technical Publishers, 2003. pp16-20(in Chinese)[Cited within: 1]
[21]
胡晓辉, 孙令强, 苗华荣, 石运庆, 陈静. 不同盐浓度对花生品种耐盐性鉴定指标的影响. 山东农业科学, 2011, (11): 35-37HuX H, SunL Q, MiaoH R, ShiY Q, ChenJ. Effects of different NaCl concentrations on indicators for evaluating salt tolerance of peanut varieties. Shand ong AgricSci, 2011, (11): 35-37 (in Chinese with English abstract)[Cited within: 1][JCR: 31.027]
[22]
TabeiY, OkadaK, HoriiE, MitsuiM, NagashimaY, SakaiT, YoshidaT, KamiyaA, FujiwaraS, TsuzukiM. Two regulatory networks mediated by light and glucose involved in glycolytic gene expression in Cyanobacteria. Plant Cell Physiol, 2012, 53: 1720-1727[Cited within: 1][JCR: 4.134]
[23]
RutterW J. Evolution of aldolase. Fed Proc, 1964, 23: 1248-1257[Cited within: 1]
[24]
LebherzH G, LeadbetterM M, BradshawR A. Isolation and characterization of the cytosolic and chloroplast forms of spinach leaf fructose diphosphate aldolase. J Biol Chem, 1984, 259: 1011-1017[Cited within: 1][JCR: 4.651]
[25]
Pelzer-ReithB, PengerA, SchnarrenbergerC. Plant aldolase: cDNA and deduced amino-acid sequences of the chloroplast and cytosol enzyme from spinach. Plant Mol Biol, 1993, 21: 331-340[Cited within: 2][JCR: 3.518]
[26]
TsutsumiK, KagayaY, HidakaS, SuzukiJ, TokairinY, HiraiT, HuD L, IshikawaK, EjiriS. Structural analysis of the chloroplastic and cytoplasmic aldolase-encoding genes implicated the occurrence of multiple loci in rice. Gene, 1994, 141: 215-220[Cited within: 2][JCR: 2.196]
[27]
KelleyP M, FreelingM. Anaerobic expression of maize fructose-1, 6-diphosphate aldolase. J Biol Chem, 1984, 259: 14180-14183[Cited within: 1][JCR: 4.651]
[28]
RussellD A, WongD M, SachsM M. The anaerobic response of soybean. Plant Physiol, 1990, 92: 401-407[Cited within: 1][JCR: 6.555]
[29]
MujerC V, RumphoM E, LinJ J, KennedyR A. Constitutive and inducible aerobic and anaerobic stress proteins in the Echinochloa complex and rice. Plant Physiol, 1993, 101: 217-226[Cited within: 1][JCR: 6.555]
[30]
AndrewsD L, MacAlpineD M, JohnsonJ R, KelleyP M, CobbB G, DrewM C. Differential induction of mRNAs for the glycolytic and ethanolic fermentative pathways by hypoxia and anoxia in maize seedlings. Plant Physiol, 1994, 106: 1575-1582[Cited within: 1][JCR: 6.555]
[31]
UmedaM, UchimiyaH. Differential transcript levels of genes associated with glycolysis and alcohol fermentation in rice plants (Oryza sativa L. ) under submergence stress. Plant Physiol, 1994, 106: 1015-1022[Cited within: 1][JCR: 6.555]
[32]
KagayaY, NakamuraH, HidakaS, EjiriS, TsutsumiK. The promoter from the rice nuclear gene encoding chloroplast aldolase confers mesophyll-specific and light-regulated expression in transgenic tobacco. Mol Gen Genet, 1995, 248: 668-674[Cited within: 1]
[33]
KamalA H, ChoK, KimD E, UozumiN, ChungK Y, LeeS Y, ChoiJ S, ChoS W, ShinC S, WooS H. Changes in physiology and protein abundance in salt-stressed wheat chloroplasts. Mol Biol Rep, 2012, 39: 9059-9074[Cited within: 1][JCR: 2.506]
[34]
SarryJ E, KuhnL, DucruixC, LafayeA, JunotC, HugouvieuxV, JourdainA, BastienO, FievetJ B, VailhenD, AmekrazB, MoulinC, EzanE, GarinJ, BourguignonJ. The early responses of Arabidopsis thaliana cells to cadmium exposure explored by protein and metabolite profiling analyses. Proteomics, 2006, 6: 2180-2198[Cited within: 1][JCR: 4.132]
[35]
张晓宁, 王昊, 曲志才, 陈火英, 叶鸣明, 沈大棱. NaCl诱导表达的盐藻果糖-1, 6-二磷酸醛缩酶基因克隆及原核表达. 复旦学报, 2002, 41: 593-595ZhangX N, WangH, QuZ C, ChenH Y, YeM M, ShenD L. Cloning and prokaryotic expression of the fructose-1, 6- diphosphate aldolase full-length cDNA of Dunaliella salina induced by NaCl. J Fudan Univ, 2002, 41: 593-595 (in Chinese with English abstract)[Cited within: 1]
[36]
PurevM, KimM K, SamdanN, YangD C. Isolation of an novel fructose-1, 6-bisphosphat aldolase gene from Codonopsis lanceolata and analysis of the response. Mol Biol, 2008, 42: 206-213[Cited within: 1][JCR: 3.518]
[37]
ZhangX N, QuZ C, WanY Z, ZhangH W, ShenD L. Application of suppression subtractive hyhridization (SSH) to cloning differentially expressed cDNA in Dunaliella salina (Chlorophyta) under hyperosmotic shock. Plant Mol Biol Rep, 2002, 20: 49-57[Cited within: 1][JCR: 5.319]
[38]
XuZ Y, KimD H, HwangI. ABA homeostasis and signaling involving multiple subcellular compartments and multiple receptors. Plant Cell Rep, 2013, 32: 807-813[Cited within: 1][JCR: 2.509]
[39]
RajjouL, BelghaziM, HuguetR, RobinC, MoreauA, JobC, JobD. Proteomic investigation of the effect of salicylic acid on Arabidopsis seed germination and establishment of early defense mechanisms. Plant Physiol, 2006, 141: 910-923[Cited within: 1][JCR: 6.555]
[40]
AbeH, UraoT, ItoT, SekiM, ShinozakiK, Yamaguchi- Shinozaki K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell, 2003, 15: 63-78[Cited within: 1][JCR: 9.251]
 , 潘丽娟
, 潘丽娟









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}