关键词:水稻(Oryza sativa L.); 叶缘白化突变体; 遗传分析; 基因定位 Genetic Analysis and Gene Mapping of a Marginal Albino Leaf Mutantmal in Rice MA Jiao, REN De-Yong, WU Guo-Chao, ZHU Xiao-Yan, MA Ling, SANG Xian-Chun, LING Ying-Hua, HE Guang-Hua* Rice Research Institute of Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops / Engineering Research Center of South Upland Agriculture, Ministry of Education, Chongqing 400716, China Fund: AbstractThe research on the color change of plant leaf is very important to clarify the structure and mechanism of photosynthetic system, such as chloroplast development and chlorophyll biosynthesis. A novel rice mutantmal (marginal albino leaf) with marginal albino leaf, was derived from the EMS-treated restorer line Jinhui 10. The mutant trait inherited steadily after several generations’ self-crossing. Themal leaf displayed albino margin and narrow blade in the whole life. Compared with the wild type,mal decreased contents of photosynthetic pigments very significantly in the whole third leaf blade, margin parts of the second and third leaves at heading stage. The observation by that transmission electronic microscopy showed cells and chloroplasts in the green part ofmalleaf developed normally, while in the albino part, the mesophyll cells were nearly hollow without obvious intact organelles and the chloroplast were fully degraded. Genetic analysis showed that the mutant trait was controlled by single recessive nuclear gene.MALwas finally mapped between SSR marker M22 and InDel marker ID27 with an interval of 171 kb on chromosome 8. These results provide a foundation for cloning and function analysis ofMAL.
Keyword:Rice (Oryza sativa L.); marginal albino leaf (mal); Genetic analysis; Gene mapping Show Figures Show Figures
图1 野生型(WT)和突变体 mal的表型特征A: 苗期野生型植株(WT); B: 苗期突变体 mal植株; C: 分蘖期野生型(WT)和突变体 mal植株; D: 分蘖期野生(WT)和突变体 mal植株叶片; E: 成熟期野生型(WT)和突变体 mal植株; F~G: 野生型(WT)和突变体 mal植株节间。Fig. 1 Plant morphology of the wild type (WT) and the mal mutantA: plants of the wild type at the seedling stage; B: plants of the mal mutant at the seedling stage; C: plants of the wild type and the mal mutant at the tillering stage; D: the leaves of the wild type and the mal mutant at the tillering stage; E: plants of the wild type and the mal mutant at the maturity stage; F-G: internode length of wide type and the mal mutant.
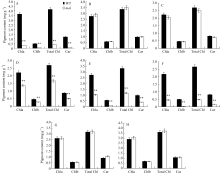
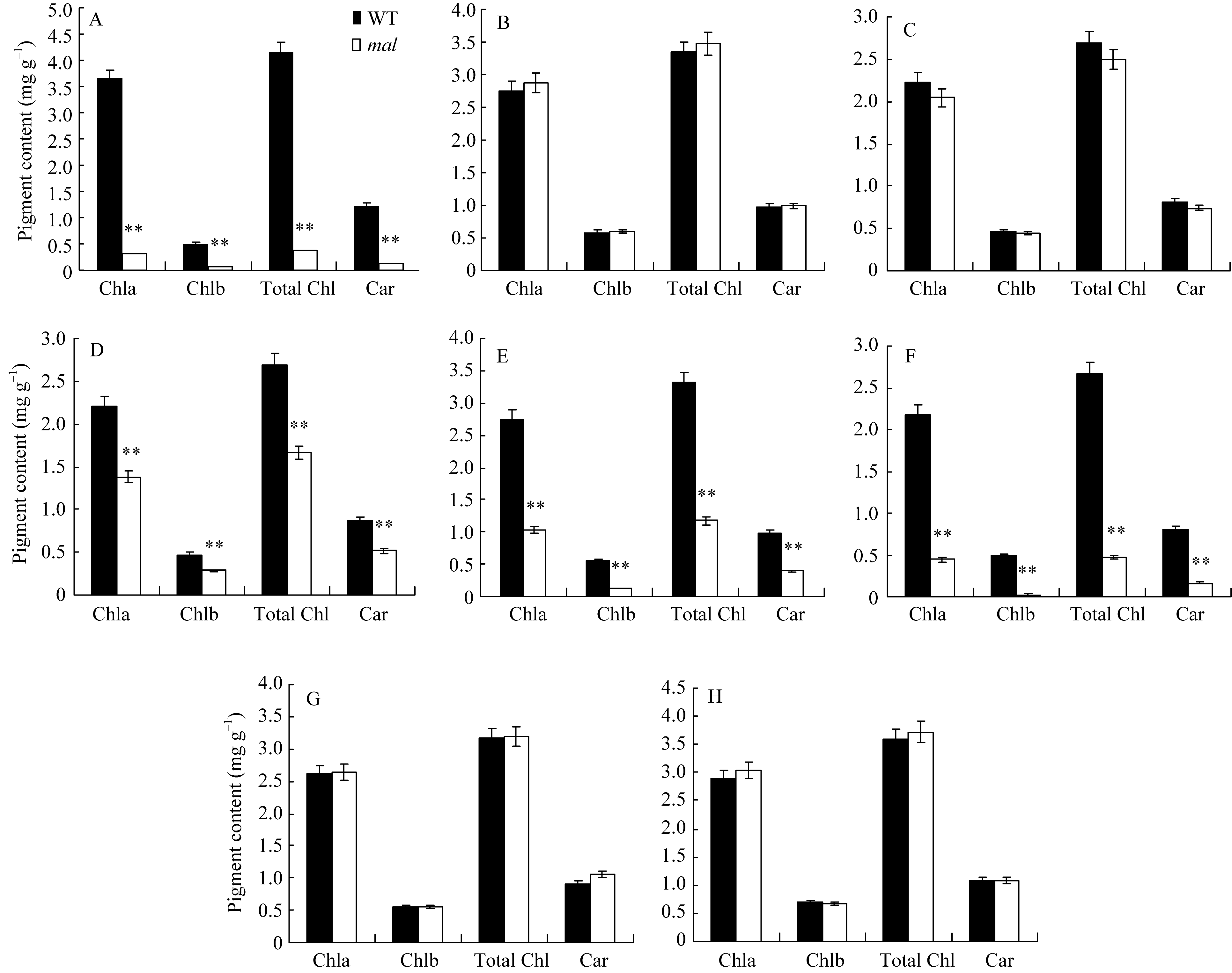
图2 野生型(WT)和 mal突变体各时期光合色素含量A: 苗期野生型(WT)和突变体 mal叶片光合色素含量; B~D: 抽穗期野生型(WT)和突变体 mal剑叶、倒二叶、倒三叶光合色素含量。E~H: 抽穗期野生型(WT)和突变体 mal倒二叶边缘、倒三叶边缘、倒二叶中部、倒三叶中部叶片光合色素含量。Chl a: 叶绿素a; Chl b: 叶绿素b; Total Chl: 叶绿素a+b的总量; Car: 类胡萝卜素。**表示在0.01水平上差异显著。Fig. 2 Photosynthetic pigments contents of wild type (WT) and malmutant at different stagesA: photosynthetic pigments contents of the malmutant and the wild type (WT) at seedling stage; B-D: photosynthetic pigments of the flag, second, and third leaf respectively in the malmutant and the wild type at heading stage. E-H: photosynthetic pigments of margin of the second and third leaf, middle green part of the second and third leaf respectively in the malmutant and the wild type at heading stage. Chl a: chlorophyll a; Chl b: chlorophyll b; Chl a+b: content of chlorophyll a and chlorophyll b; Car: carotenoids. ** Significantly different at P<0.01 by t-test as determined by t-test.
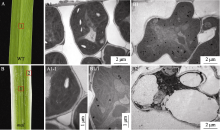
图3 野生型(WT)和突变体 mal的叶肉细胞及叶绿体的超微结构观察A: 野生型叶片; B: mal突变体叶片; A1: 野生型叶肉细胞结构; B1: mal叶片绿色部分叶肉细胞结构; A1-1: 野生型叶绿体结构; B1-1: mal叶片绿色部分的叶绿体结构; B2: mal叶片白色部分的叶肉细胞结构。Fig. 3 Ultrastructures of chloroplasts in the mesophyll cells of the wild-type (WT) and mal mutantA: leaf of wild-type; B: Leaf of mal; A1: structure of wild-type mesophyll cell; B1: mesophyll cell structure in green sectors of mal leaves; A1-1: chloroplast structure in wild-type; B1-1: chloroplast structure in green sectors of malleaves; B2: mesophyll cell structure in white sectors of mal leaves.
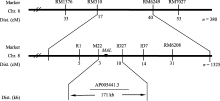
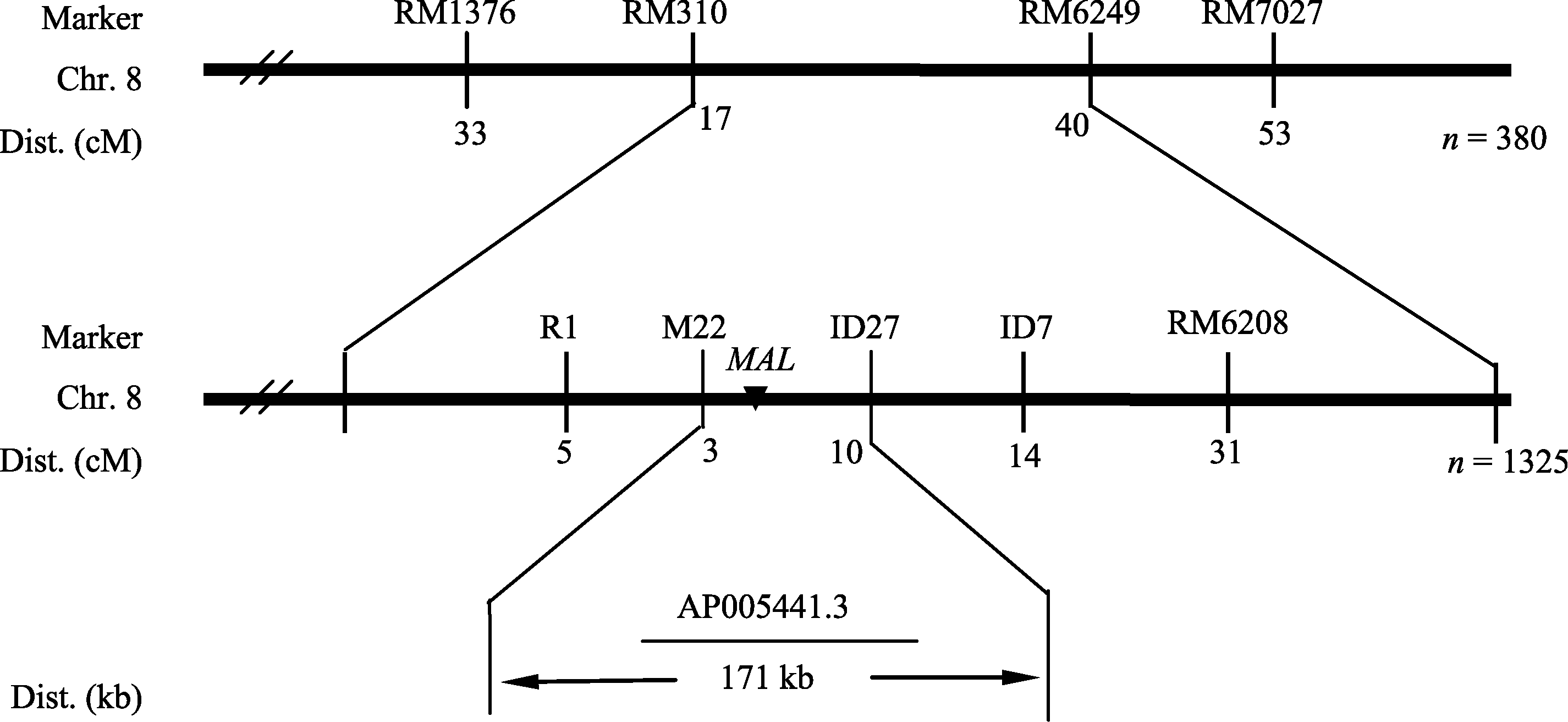
4 结论 mal突变体在整个生育期叶片均呈边缘白化, 光合色素含量和净光合速率显著降低, 叶片变窄, 株高、有效穗数、每穗粒数、每穗实粒数与千粒重极显著降低。 mal突变体叶片绿色部分细胞及其叶绿体发育完全, 而白化部分细胞结构严重缺陷, 叶绿体结构降解。该突变性状受1对隐性核基因控制, 该基因被定位在第8染色体SSR标记M22和InDel标记ID27之间, 遗传距离分别为0.22 cM和0.73 cM, 物理距离为171 kb, 是一个新的叶色突变基因。 ** The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
KlimyukV I, Persello-CartieauxF, HavauxM, Contard-DavidP, SchuenemannD, MeiherhoffK, GouetP, Jones J D G, HoffmancN E, LaurentNussaume. A chromodomain protein encoded by the Arabidopsis CAO gene is a plant-specific component of the chloroplast signal recognition particle pathway that is involved in LHCP targeting. Plant Cell, 1999, 1: 87-99[本文引用:1][JCR: 9.251]
[2]
ChenG, BiY R, LiN. EGY1 encodes a membrane-associated and ATP-independent metalloprotease that is required for chloroplast development. Plant J, 2005, 41: 364-375[本文引用:2][JCR: 6.582]
[3]
ChenT, ZhangY, ZhaoL, ZhuZ, LinJ, ZhangS B, WangC L. Physiological character and gene mapping in a new green- revertible albino mutant in rice. Genet Genomics, 2007, 34: 331-338[本文引用:1][JCR: 2.397]
[4]
MotohashiR, ItoT, KobayashiM, TajiT, NagataN, AsamiT, YoshidaS, Yamaguchi-ShinozakiK, ShinozakiK. Functional analysis of the 37 kDa inner envelope membrane polypeptide in chloroplast biogenesis using a Ds-tagged Arabidopsis pale green mutant. Plant J, 2003, 34: 719-731[本文引用:1][JCR: 6.582]
[5]
SugimotoH, KusumiK, TozawaY, YazakiJ, KishimotoN, KikuchiS, IbaK. The virescent-2 mutation inhibits translation of plastid transcripts for the plastid genetic system at an early stage of chloroplast differentiation. Plant Cell Physiol, 2004, 45: 958-996[本文引用:1][JCR: 4.134]
[6]
IhnatowiczA, PesaresiP, VarottoC, RichlyE, SchneiderA, JahnsP, SalaminiF, LeisterD. Mutants for photosystem I subunit D of Arabidopsis thaliana: effects on photosynthesis, photosystem I stability and expression of nuclear genes for chloroplast functions. Plant J, 2004, 37: 839-852[本文引用:1][JCR: 6.582]
[7]
NagataN, TanakaR, SatohS, TanakaA. Identification of a vinyl reductase gene for chlorophyll synthesis in Arabidopsis thaliana and implications for the evolution of Prochlorococcus species. Plant Cell, 2005, 17: 233-240[本文引用:1][JCR: 9.251]
[8]
JungK H, HurJ, RyuC H, ChoiY. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol, 2003, 44: 463-472[本文引用:2][JCR: 4.134]
[9]
LarkinR M, AlonsoJ M, EckerJ R, ChoryJ. GUN4, a regulator of chlorophyll synthesis and intracellular signaling. Science, 2003, 299: 902-906[本文引用:2]
[10]
董凤高, 朱旭东, 熊振民, 程式华, 孙宗修, 闵绍楷. 以淡绿叶为标记的籼型一温敏核不育系M2S的选育. 中国水稻科学, 1995, 9: 65-70DongF G, ZhuX D, XiongZ M, ChengS H, SunZ X, MinS K. Breeding of a photo-thermoperiod sensitive genic male sterile indica rice with a pale-green-leaf marker. Chin J Rice Sci, 1995, 9: 65-70 (in Chinese with English abstract)[本文引用:2][CJCR: 1.494]
[11]
吴自明, 张欣, 万建民. 水稻黄绿叶基因的克隆及应用. 生命科学, 2007, 12: 614-615WuZ M, ZhangX, WanJ M. Cloning and application of yellow-green leaf genes in rice. Chin Bull Life Sci, 2007, 12: 614-615 (in Chinese)[本文引用:2][CJCR: 0.577]
[12]
IwataN, OmuraT. Studies on the trisomics in rice plants (Oryza sativa L. ): III. Relation between trisomics and genetic linkage groups. Jpn J Breed, 1975, 25: 363-368[本文引用:2]
[13]
ZhangH, LiJ J, YooJ H, YooS C, ChoS H, KohH J, SeoH S, PaekN C. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol, 2006, 62: 325-337[本文引用:1][JCR: 3.518]
[14]
LeeS, KimJ H, YooE S, LeeC H, HirochikaH, AnG. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol Biol, 2005, 57: 805-818[本文引用:1][JCR: 3.518]
[15]
WuZ M, ZhangX, HeB, DiaoL P, ShengS L, WangJ L, GuoX P, SuN, WangL F, JiangL, WangC M, ZhaiF Q, WanJ M. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:2][JCR: 6.555]
[16]
YooS C, ChoS H, SugimotoH, LiJ, KusumiK, KohH J, KohI, PaekN C. Rice virescent3 and stripe1 encoding the large and small subunits of ribonucleotide reductase are required for chloroplast biogenesis during early leaf development. Plant Physiol, 2009, 150: 388-401[本文引用:2][JCR: 6.555]
[17]
SugimotoH, KusumiK, NoguchiK, YanoM, YoshimuraA, IbaK. The rice nuclear gene, VIRESCENT 2, is essential for chloroplast development and encodes a novel type of guanylate kinase targeted to plastids and mitochondria. Plant J, 2007, 52: 512-527[本文引用:2][JCR: 6.582]
[18]
KusabaM, ItoH, MoritaR, IidaS, SatoY, FujimotoM, KawasakiS, TanakaR, HirochikaH, NishimuraM, TanakaA. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell, 2007, 19: 1362-1375[本文引用:1][JCR: 9.251]
[19]
ParkS Y, YuJ W, ParkJ S, LiJ, YooS C, LeeN Y, LeeS K, JeongS W, SeoH S, KohH J, JeonJ S, ParkY I, PaekN C. The senescence-induced stay-green protein regulates chlorophyll degradation. Plant Cell, 2007, 19: 1649-1664[本文引用:1][JCR: 9.251]
[20]
WangP R, GaoJ X, WanC M, ZhangF T, XuZ J, HuangX Q, SunX Q, DengX J. Divinyl chlorophyll(ide) α can be converted to monovinyl chlorophyll(ide) α by a divinyl reductase in rice. Plant Physiol, 2010, 153: 994-1003[本文引用:1][JCR: 6.555]
[21]
SakurabaY, RahmanM L, ChoS H, KimY S, KohH J, YooS C, PaekN C. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J, 2013, 74: 122-133[本文引用:1][JCR: 6.582]
[22]
DongH, FeiG L, WuC Y, WuF Q, SunY Y, ChenM J, RenY L, ZhouK N, ChengZ J, WangJ L, JiangL, ZhangX, GuoX P, LeiC L, SuN, WangH Y, WanJ M. A rice virescent-yellow leaf mutant reveals new insights into the role and assembly of plastid caseinolytic protease in higher plants. Plant Physiol, 2013, 162: 1867-1880[本文引用:1][JCR: 6.555]
[23]
LichtenthalerH K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Method Enzymol, 1987, 48: 350-382[本文引用:1][JCR: 2.002]
[24]
张守仁. 叶绿素荧光动力学参数的意义及讨论. 植物学通报, 1999, 16: 444-448ZhangS R. A discussion on chlorophyll fluorescence kinetics parameters and their significance. Chin Bull Bot, 1999, 16: 444-448 (in Chinese with English abstract)[本文引用:1]
[25]
何瑞峰, 丁毅, 余金洪, 祖明生. 水稻温敏叶绿素突变体叶片超微结构的研究. 武汉植物学研究, 2001, 19: 1-5HeR F, DingY, YuJ H, ZuM S. Study on leaf ultrastructure of the thermo-sensitive chlorophyll deficient mutant in rice. J Wuhan Bot Res, 2001, 19: 1-5 (in Chinese with English abstract)[本文引用:1]
[26]
MichelmoreR W, ParanI, KesseliR V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1][JCR: 9.737]
[27]
MurrayM G, ThompsonW F. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4325[本文引用:1]
[28]
桑贤春, 何光华, 张毅, 杨正林, 裴炎. 水稻PCR扩增模板的快速制备. 遗传, 2003, 25: 705-707SangX C, HeG H, ZhangY, YangZ L, PeiY. The simple gain of templates of rice genomes DNA for PCR. Hereditas (Beijing), 2003, 25: 705-707 (in Chinese with English abstract)[本文引用:1]
[29]
PanaudO, ChenX, McCouchS R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L. ). Mol Gen Genet, 1996, 252: 597-607[本文引用:1]
[30]
徐培洲, 李云, 袁澍, 张红宇, 彭海, 林宏辉, 汪旭东, 吴先军. 叶绿素缺乏水稻突变体中光系统蛋白和叶绿素合成特性的研究. 中国农业科学, 2006, 39: 299-305XuP Z, LiY, YuanS, ZhangH Y, PengH, LinH H, WangX D, WuX J. Studies of photosystem complexes and chlorophyll synthesis in chlorophyll-deficient rice mutant W1. Sci Agric Sin, 2006, 39: 299-305 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[31]
ZhaoY, DiL F, YangS H, LiS C, ZangY Z. Chloroplast composition and structural differences in a chlorophyll reduced mutant of oilseed rape seedlings. Acta Bot Sin, 2001, 43: 877-880[本文引用:1][JCR: 0.599]
[32]
HuangX Q, WangP R, ZhaoH X, DengX J. Genetic analysis and molecular mapping of a novel chlorophyll deficit mutant gene in rice. Rice Sci, 2008, 15: 7-12[本文引用:1][CJCR: 0.3579]
[33]
LiuW Z, FuY P, HuG C, SiH M, ZhuL, WuC, SunZ X. Identification and fine mapping of a thermo sensitive chlorophyll deficient mutant in rice (Oryza sativa L. ). Planta, 2007, 226: 785-795[本文引用:1][JCR: 3.347]
[34]
IwataN, SatohH, OmuraT. Linkage analysis by use of trisomics in rice (Oryza sativa L. ): IV. Linkage groups locating on chromosomes 2 and 10. Jpn J Breed, 1981, 31: 66-67[本文引用:1]
[35]
程世超, 刘合芹, 翟国伟, 冯世座, 赵辉, 汪得凯, 陶跃之. 水稻白化致死突变体abl4的鉴定和基因定位. 中国水稻科学, 2013, 27: 240-246ChengS C, LiuH Q, ZhaiG W, FengS Z, ZhaoH, WangD K, TaoY Z. Genetic analysis and gene mapping of an albino lethal mutant in rice. Chin J Rice Sci, 2013, 27: 240-246 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[36]
余庆波, 江华, 米华玲, 周根余, 杨仲南. 水稻白化突变体alb21生理特性和基因定位. 上海师范大学学报, 2005, 34(1): 70-75YuQ B, JiangH, MiH L, ZhouG Y, YangZ N. Physiological property and gene mapping of an albino mutant alb21 in rice. J Shanghai Norm Univ, 2005, 34(1): 70-75 (in Chinese)[本文引用:1]
[37]
李育红, 王宝和, 戴正元, 李爱宏, 赵步洪, 左示敏, 陈忠祥, 张洪熙, 潘学彪. 水稻叶色突变体及其基因定位、克隆的研究进展. 江苏农业科学, 2011, 39(2): 34-39LiY H, WangB H, DaiZ Y, LiA H, ZhaoB H, ZuoS M, ChenZ X, ZhangH X, PanX B. The research advances of gene mapping and cloning of leaf color mutants in rice. Jiangsu Agric Sci, 2011, 39(2): 34-39 (in Chinese)[本文引用:1]
[38]
ParksB M, QuailP H. Phytochrome-deficient hyl and hy2 long hypocotyls mutants of Arabidopsis are defective in phytochrome chromophore biosynthesis. Plant Cell, 1991: 1177-1186[本文引用:2][JCR: 9.251]
[39]
朱丽, 刘文真, 吴超, 栾维江, 傅亚萍, 胡国成, 斯华敏, 孙宗修. 水稻着丝粒附近一个淡绿叶突变相关基因的定位分析. 中国水稻科学, 2007, 21: 228-234ZhuL, LiuW Z, WuC, LuanW J, FuY P, HuG C, SiH M, SunZ X. Identification and fine mapping of a gene related to pale green leaf near centromere region in rice (Oryza sativa L. ). Chin J Rice Sci, 2007, 21: 228-234 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[40]
XiaJ C, WangY P, MaB T, YinZ Q, HaoM, KongD W, LiS G. Ultrastructure and gene mapping of the albino mutant al12 in rice (Oryza sativa L. ). Acta Genet Sin, 2006, 33: 1112-1119[本文引用:1]
[41]
SiddappaK, VasudevK L, GanigerB S, RathodR, DevarK V. Report of albino seedlings in Pongamia pinnata. Karnataka J Agric Sci, 2004, 17: 884-885[本文引用:1]
[42]
ChenT, ZhangY D, ZhaoL, ZhuZ, LinJ, ZhangS B, WangC L. Physiological character and gene mapping in a new green-revertible albino mutant in rice. J Genet Genomics, 2007, 34: 331-338[本文引用:1][JCR: 2.076][CJCR: 1.323]
[43]
ChenT, ZhangY D, ZhaoL, ZhuZ, LinJ, ZhangS B, WangC L. Fine mapping and cand idate gene analysis of a green- revertible albino gene gra(t) in rice. J Genet Genomics, 2009, 36: 117-123[本文引用:1][JCR: 2.076][CJCR: 1.323]
[44]
LanT, WangB, LingQ P, XuC H, TongZ J, LiangK J, DuanY L, JinJ, WuW R. Fine mapping of cisc(t), a gene for cold-induced seedling chlorosis, and identification of its cand idate in rice. Chin Sci Bull, 2010, 55: 3149-3153[本文引用:1][CJCR: 0.95]





{kind=link}
{kind=link}
{kind=link}
{kind=link}