关键词:簇毛麦; 高分子量谷蛋白亚基; 原核表达; 掺粉实验 Isolation, Characterization and Farinograph Analysis of Novel HMW-GSs fromDasypyrum villosum YANG Hua1, GAO Xiang1,2,*, CHEN Qi-Jiao1, ZHAO Wan-Chun1,*, DONG Jian1,2, LI Xiao-Yan1 1 College of Agronomy, Northwest A&F University, Yangling 712100, China
2 Wheat Engineering Research Center of Shaanxi Province / New Varieties Cultivation of Wheat Engineering Research Center of Shaanxi Province, Yangling 712100, China
Fund: AbstractDasypyrum villosum carrying many novel HMW-GS alleles is an important genetic resource for wheat protein improvement. In this study, we isolated six HMW-GS genes fromD. villosum TA10220 (1.0-1.7 kb, GenBank accession numbers: KF887414-KF887419), which were substantially smaller than those from common wheat, using a pair of specific primers. An in-frame stop codon was found in the coding sequences of KF887418 and KF887419 and thus these genes might be pseudogenes. The comprehensive analysis of deduced amino acid sequence, and phylogenetic and evolutionary analyses of full sequence, N- and C-terminal domains revealed that KF887414 was closely related to y-type HMW-GS, but KF887415-KF887419 had structural characteristics of both x- and y-types. DNA fragments of KF887414-KF887417 were subcloned into the pEASY-E2 expression vector and expressed inEscherichia coli Rosetta-gami B(DE3) cell under IPTG induction. The four genes were successfully expressed inE. coli system according to SDS-PAGE analysis (both the expressed protein and HMW-GS isolated from seed) and western-blotting assay. The fusion protein was purified and recovered by His-Trap affinity chromatography and low temperature cryodesiccation, and then integrated into the control flour by using a 4 g Micro-dough LAB Farinograph. Results showed that the four HMW-GSs originated fromD. villosum had positive effects on dough quality property.
Keyword:Dasypyrum villosum; High-molecular-weight glutenin subunits (HMW-GS); Prokaryotic expression; Farinograph Show Figures Show Figures
表1 核苷酸序列和氨基酸序列特征 Table 1 Basic characteristics of nucleotide sequence and amino acid sequence
登录号1) Accession number1)
核苷酸序列 Nucleotide sequence
推导产物 Deduced amino acid sequence
融合表达蛋白的分子量3) Molecular weight of fusion protein product (kD)3)
序列长度 Length (bp)
开放阅读框位置 Position of ORF
氨基酸个数2) Size of putative protein2)
分子量2) Molecular weight (kD)2)
KF887414
1665
61-1665 bp
534
57.308
57.355
KF887415
1296
61-1296 bp
411
45.034
45.081
KF887416
1152
61-1152 bp
363
39.809
39.841
KF887417
1152
61-1152 bp
363
39.866
39.898
KF887418
1539
61-1539 bp
492
53.729
—
KF887419
1377
61-1377 bp
438
47.824
—
1)下画线表示假基因; 带下画线的序列为假基因;2)推导的氨基酸序列包括信号肽;3)融合表达的蛋白没有信号肽, 但包含有2.101 kD的His-Tag标签。 推导产物含信号肽。 1) The pseudogenes are underlined;2) The deduced products contain signal peptides;3) The expressed proteins without signal peptides contain His-Tag of 2.101 kD.
表1 核苷酸序列和氨基酸序列特征 Table 1 Basic characteristics of nucleotide sequence and amino acid sequence
图2 KF887414、KF887415、KF887416、KF887417、KF887418和KF887419推导氨基酸序列分析Fig. 2 Deduced amino acid sequences of KF887414, KF887415, KF887416, KF887417, KF887418, and KF887419
表2 Table 2 表2(Table 2)
表2 不同来源的HMW-GS的主要结构特征的比较 Table 2 A summary of properties of the primary structure of HMW-GS from Dasypyrum villosumin comparison with some HMW-GS from common wheat and wheat-related grass
表2 不同来源的HMW-GS的主要结构特征的比较 Table 2 A summary of properties of the primary structure of HMW-GS from Dasypyrum villosumin comparison with some HMW-GS from common wheat and wheat-related grass
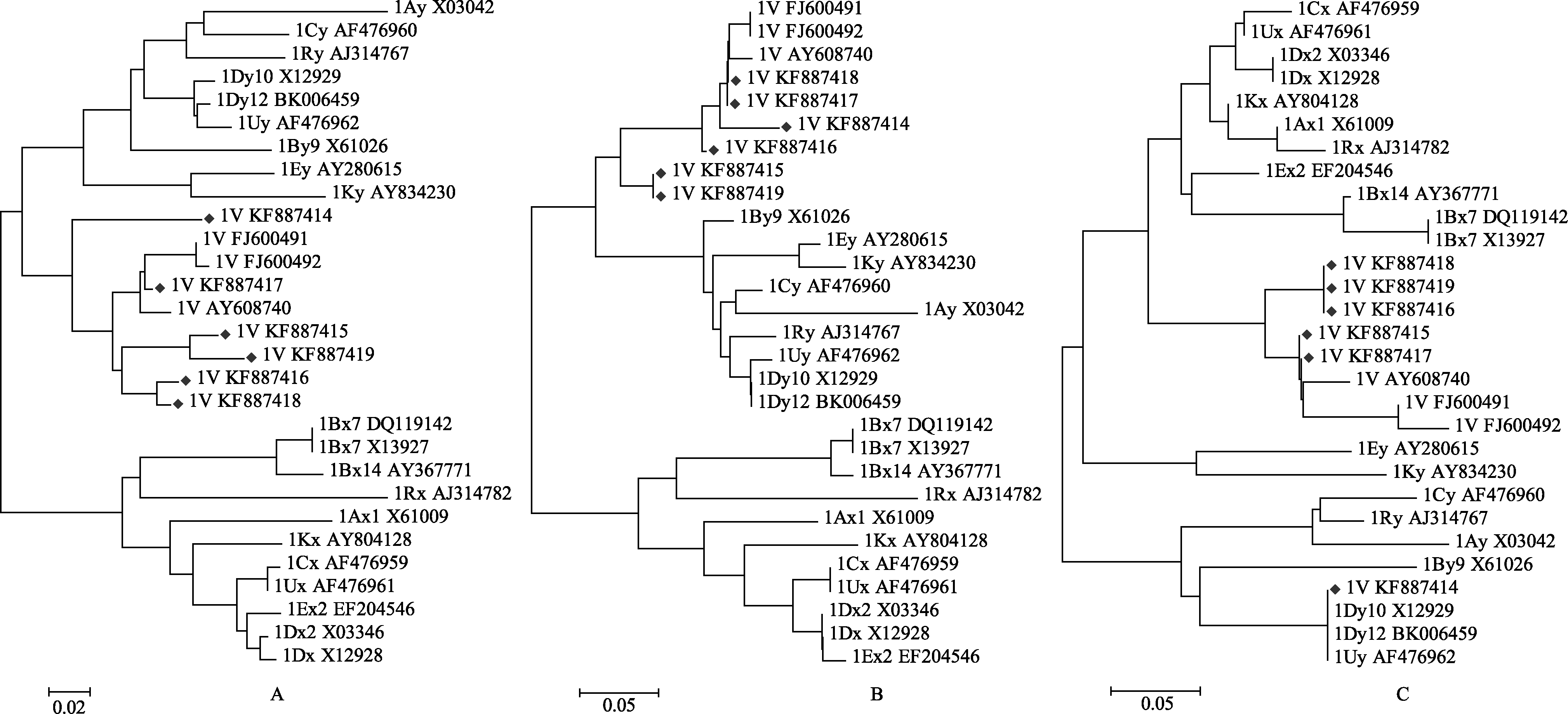
图3 簇毛麦HMW-GS编码的蛋白序列系统演化分析(Neighbor-joining进化树)Fig. 3 Phylogenetic analysis of the deduced amino acid sequence of HMW-GS from Dasypyrum villosum (Neighbor-joining tree)
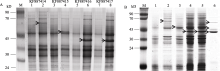
图4 KF887414、KF887415、KF887416和KF887417基因的诱导表达产物和种子HMW-GS的SDS-PAGE分析M: 蛋白质分子质量标准Marker II。A图中, 1、3、5和7泳道为未诱导重组质粒表达产物; 2、4、6和8泳道为经IPTG诱导后的重组质粒的表达产物; 箭头示基因的表达产物。B图中, 泳道1为未诱导重组质粒表达产物; 2~5泳道为经IPTG诱导后的重组质粒的表达产物, 依次是 KF887414、KF887415、KF887416和KF887417基因; 泳道6为种子提取的HMW-GS; 箭头示目的蛋白。Fig. 4 Analyses of expressed proteins of genes from KF887414 to KF887417 and comparison with seed HMW-GS using SDS-PAGEM: Protein ladder marker II. In panel A, lanes 1, 3, 5, and 7 show proteins of recombinant plasmid before induction and lanes 2, 4, 6, and 8 show proteins of recombinant plasmid after adding IPTG; the arrows show expression products. In panel B, lane 1 shows the protein of recombinant plasmid before induction; lanes 2, 3, 4, and 5 show proteins of recombinant plasmid KF887414, KF887415, KF887416, and KF887417 after adding IPTG, respectively; lane 6 shows HMW-GS extracted from Dasypyrum villosum seed; the arrows show target proteins.
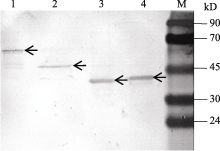
图5 KF887414、KF887415、KF887416和KF887417基因的诱导表达产物Western blot检测M: 蛋白质分子质量标准Marker II。1、2、3和4泳道分别为KF887414、KF887415、KF887416和KF887417基因经IPTG诱导后的表达产物; 箭头示基因的表达产物。Fig. 5 Western blotting assay of expressed proteins of genes from KF887414 to KF887417M: Protein ladder marker II; lanes 1, 2, 3, and 4 show proteins of recombinant plasmid KF887414, KF887415, KF887416, and KF887417 after adding IPTG, respectively; the arrows show target proteins.
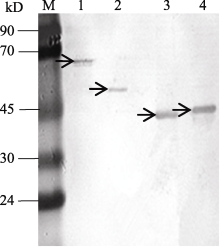
图6 KF887414、KF887415、KF887416和KF887417基因的诱导表达产物柱纯化M: 蛋白质分子质量标准Marker II。1、2、3和4泳道分别为KF887414、KF887415、KF887416和KF887417基因经IPTG诱导后的表达产物; 箭头示基因的表达产物。Fig. 6 Analyses of purification of expressed proteins of genes from KF887414 to KF887417 using SDS-PAGEM: Protein ladder marker II; lanes 1, 2, 3, and 4 show proteins of recombinant plasmid KF887414, KF887415, KF887416, and KF887417 after adding IPTG, respectively; the arrows show target proteins.
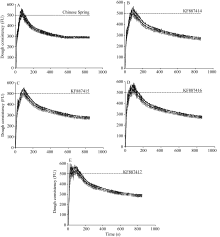
图7 簇毛麦来源的HMW-GS对粉质曲线的影响A: 中国春添加DTT、KIO3; B: 中国春添加DTT、KIO3和KF887414; C: 中国春添加DTT、KIO3和KF887415; D: 中国春添加DTT、KIO3和KF887416; E: 中国春添加DTT、KIO3和KF887417。Fig. 7 Effects of HMW-GS from Dasypyrum villosumon farinograph parametersA: Chinese Spring with DTT and KIO3; B: Chinese Spring with DTT, KIO3,and KF887414; C: Chinese Spring with DTT, KIO3,and KF887415; D: Chinese Spring with DTT, KIO3,and KF887416; E: Chinese Spring with DTT, KIO3,and KF887417.
表3 Table 3 表3(Table 3)
表3 纯化蛋白亚基的粉质参数 Table 3 Farinograph parameters of purified HMW-GS from Dasypyrum villosum
添加物 Additive
形成时间 DvT (min)
稳定时间 ST (min)
弱化度 DS (FU)
机械耐力系数 MTI (FU)
及线时间 AT (min)
离线时间 DT (min)
粉质质量参数 FQN (mm)
DTT, KIO3
1.2 C
0.9 C
174.9 A
198 A
0.9 B
1.7 A
4.83 E
DTT, KIO3 KF887414
1.6 A
1.2 B
154.9 C
170 C
1.1 A
2.3 C
7.67 A
DTT, KIO3 KF887415
1.6 A
1.2 B
159.9 B
180 B
1.1 A
2.3 C
6.67 D
DTT, KIO3 KF887416
1.5 B
1.2 B
174.9 A
185 B
0.9 B
2.2 B
7.50 B
DTT, KIO3 KF887417
1.5 B
1.5 A
174.9 A
185 B
0.7 C
2.2 B
7.17 C
表3 纯化蛋白亚基的粉质参数 Table 3 Farinograph parameters of purified HMW-GS from Dasypyrum villosum
表中数据为3次试验的平均值, 均值后大写字母表示0.01差异显著水平。 Data are the average of triplicated micro-mixing experiments. Means marked with different letters are significantly different at P < 0.01. DvT: development times; ST: stability time; DS: degree of time; MTI: mixing tolerance index; AT: arrival time; DT: departure time; FQN: farinograph quality number. 图8 Fig. 8
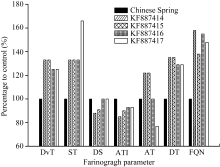
图8 添加KF887414、KF887415、KF887416和KF887417基因表达蛋白的中国春与中国春(对照)粉质参数的比较DvT: 形成时间; ST: 稳定时间; DS: 弱化度; MTI: 机械耐力系数; AT: 及线时间; DT: 离线时间; FQN: 粉质质量参数。Fig. 8 Comparison of farinogragh parameters between Chinese Spring (control) and Chinese Spring with expressed proteins of KF887414, KF887415, KF887416, and KF887417DvT: development time; ST: stability time; DS: degree of time; MTI: mixing tolerance index; AT: arrival time; DT: departure time; FQN: farinograph quality number.
WrigleyC W. Biopolymers-Giant proteins with flour power. Nature, 1996, 381: 738-739[本文引用:1][JCR: 38.597]
[2]
PayneP I. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu Rev Plant Phys, 1987, 38: 141-153[本文引用:5]
[3]
ShewryP R, HalfordN G, TathamA S. High molecular weight subunits of wheat glutenin. J Cereal Sci, 1992, 15: 105-120[本文引用:6][JCR: 2.088]
[4]
LawrenceG J, ShepherdK W. Variation in glutenin protein subunits of wheat. Aust J Biol Sci, 1980, 33: 221-233[本文引用:1]
[5]
LawrenceG J, ShepherdK W. Inheritance of glutenin protein subunits of wheat. Theor Appl Genet, 1981, 60: 333-337[本文引用:1][JCR: 3.658]
[6]
PayneP I, LawC N, MuddE E. Control by homoeologous group 1 chromosomes of the high-molecular-weight subunits of glutenin, a major protein of wheat endosperm. Theor Appl Genet, 1980, 58: 113-120[本文引用:1][JCR: 3.658]
[7]
PayneP I, HoltL M, LawC N. Structural and genetical studies on the high- molecular-weight subunits of wheat glutenin. Theor Appl Genet, 1982, 63: 129-138[本文引用:1][JCR: 3.658]
[8]
PayneP I, NightingaleM A, KrattigerA F, HoltL M. The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J Sci Food Agric, 1987, 40: 51-65[本文引用:1][JCR: 1.759]
[9]
MackieA M, LagudahE S, SharpP J, Lafiand raD. Molecular and biochemical characterization of HMW glutenin subunits from T. tauschii and the D genome of hexaploid wheat. J Cereal Sci, 1996, 23: 213-225[本文引用:2][JCR: 2.088]
[10]
HarberdN P, BartelsD, ThompsonR D. DNA restriction- fragment variation in the gene family encoding high molecular weight (HMW) glutenin subunits of wheat. Biochem Genet, 1986, 24: 579-596[本文引用:1][JCR: 0.938]
[11]
ShewryP R, HalfordN G, TathamA S. The high molecular weight subunits of wheat, barley and rye: genetics, molecular biology, chemistry and role in wheat gluten structure and functionality. Oxford Surv Plant Mol Cell Biol, 1989, 6: 163-219[本文引用:2]
[12]
DovidioR, MasciS, PorcedduE. Development of a set of oligonucleotide primers specific for genes at the Glu-1 complex loci of wheat. Theor Appl Genet, 1995, 91: 189-194[本文引用:2][JCR: 3.658]
[13]
GradzielewskaA. The genus Dasypyrum: 1. The taxonomy and relationships within Dasypyrum and with Triticeae species. Euphytica, 2006, 152: 429-440[本文引用:2][JCR: 1.643]
[14]
GradzielewskaA. The genus Dasypyprum: 2. Dasypyrum villosum: a wild species used in wheat improvement. Euphytica, 2006, 152: 441-454[本文引用:2][JCR: 1.643]
[15]
DePace C, SnidaroD, CiaffiM, VittoriD, CiofoA, CenciA, TanzarellaO A, QualsetC O, Scarascia Mugnozza G T. Introgression of Dasypyrum villosum chromatin into common wheat improves grain protein quality. Euphytica, 2001, 117: 67-75[本文引用:2][JCR: 1.643]
[16]
ZhaoW C, QiL L, GaoX, ZhangG S, DongJ, ChenQ J, BerndF, BikramS G. Development and characterization of two new Triticum aestivum-Dasypyrum villosum Robertsonian translocation lines T1DS·1V#3L and T1DL·1V#3S and their effect on grain quality. Euphytica, 2010, 175: 343-350[本文引用:2][JCR: 1.643]
[17]
董剑, 杨华, 赵万春, 李晓燕, 陈其皎, 高翔. 普通小麦中国春-簇毛麦异位系T1DL·1VS和T1DS·1VL的农艺和品质特性. 作物学报, 2013, 39: 1386-1390DongJ, YangH, ZhaoW C, LiX Y, ChenQ J, GaoX. Agronomic traits and grain quality of Chinese Spring-Dasypyrum villosum translocation lines T1DL·1VS and T1DS·1VL. Acta Agron Sin, 2013, 39: 1386-1390 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[18]
MonteboveL, DePace C, JanC C, Scarascia Mugnozza G T, QualseC O. Chromosomal location of isozyme and seed storage protein genes in Dasypyrum villosum (L. ) Cand argy. Theor Appl Genet, 1987, 73: 836-845[本文引用:2][JCR: 3.658]
[19]
ShewryP R, ParmarC, PappinD C. Characterization and genetic control of the prolamins of Haynaldia villosa: relationship to cultivated species of the Triticeae (rye, wheat, and barley). Biochem Genet, 1987, 25: 309-325[本文引用:1][JCR: 0.938]
[20]
张瑞奇. 簇毛麦籽粒硬度基因和贮藏蛋白基因的染色体定位及易位系选育. 南京农业大学博士学位论文, 2010ZhangR Q. Chromosome Location of Genes for Hardness Locus and Storage Proteins in H. villosa and Creation the Translocation Lines. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2010 (in Chinese with English abstract)[本文引用:3]
[21]
庞玉辉, 陈新宏, 赵继新, 武军, 程雪妮, 刘淑会, 杨群慧, 杜万里, 陈林刚. 簇毛麦HMW-GS及其启动子基因的克隆与序列分析. 西北植物学报, 2009, 29: 859-866PangY H, ChenX H, ZhaoJ X, WuJ, ChengX N, LiuS H, YangQ H, DuW L, ChenL G. Cloning and sequence analysis of the HMW-GS gene and its promoter from Dasypyrum villosum. Acta Bot Boreali-Occident Sin, 2009, 29: 859-866 (in Chinese with English abstract)[本文引用:4][CJCR: 1.321]
[22]
陈凡国, 朱翔宇, 夏光敏. 簇毛麦中一种新型高分子量麦谷蛋白亚基基因序列的研究. 西北植物学报, 2005, 25: 1410-1414ChenF G, ZhuX Y, XiaG M. Sequence of a new HMW glutenin subunit gene in Haynaldia villosa. Acta Bot Boreali-Occident Sin, 2005, 25: 1410-1414 (in Chinese with English abstract)[本文引用:3][CJCR: 1.321]
[23]
刘守斌. 簇毛麦染色体分子标记的筛选及其高分子量谷蛋白亚基基因的克隆. 中国农业大学博士学位论文, 2002LiuS B. The Characterization of V-Genome Specific Molecular Marker and Cloning of High-Molecular-Weight Glutenin Subunit Genes in Haynaldia villosa. PhD Dissertation of China Agricultural University, Beijing, China, 2002 (in Chinese with English abstract)[本文引用:3]
[24]
陈晓燕. 簇毛麦储藏蛋白基因的克隆及小麦的抗白粉病育种. 山东大学硕士学位论文, 2009ChenX Y. Glutenin and Gliadin Genes Cloning from Dasypyrum villosum and Powdery Mildew Resistance Breeding in Wheat. MS Thesis of Shand ong University, Tai’an, Shand ong, China, 2009 (in Chinese with English abstract)[本文引用:3]
VerbruggenI M, VeraverbekeW S, Vand ammeA, DelcourA. Simultaneous isolation of wheat high molecular weight and low molecular weight glutenin subunits. J Cereal Sci, 1998, 28: 25-32[本文引用:2][JCR: 2.088]
[27]
MurrayM G, ThompsonW F. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4325[本文引用:1]
[28]
XuH, WangR J, ShenX, ZhaoY L, SunG L, ZhaoH X, GuoA G. Functional properties of a new low-molecular-weight glutenin subunit gene from a bread wheat cultivar. Theor Appl Genet, 2006, 113: 1295-1303[本文引用:1][JCR: 3.658]
[29]
ChenF G, ZhaoF, LiuR K, XiaG M. Functional properties of two low-molecular-weight glutenin subunits carrying additional cysteine residues from hybrid introgression line II-12 derived from Triticum aestivum and Agropyron elongatum. Food Chem, 2011, 127: 1773-1776[本文引用:1][JCR: 3.334]
[30]
FordeJ, MalpicaJ M, HalfordN G, ShewryP R, AndersonO D, GreeneF C, MiflinB J. The nucleotide sequence of a HMW glutenin subunit gene located on chromosome 1A of wheat (Triticum aestivum L. ). Nucl Acids Res, 1985, 13: 6817-6832[本文引用:1]
[31]
DeBustos A, RubioP, JouveN. Molecular characterisation of the inactive allele of the gene Glu-A1 and the development of a set of AS-PCR markers for HMW glutenins of wheat. Theor Appl Genet, 2000, 100: 1085-1094[本文引用:1][JCR: 3.658]
[32]
WanY, WangD, ShewryP, HalfordN. Isolation and characterization of five novel high molecular weight subunit of glutenin genes from Triticum timopheevi and Aegilops cylindrica. Theor Appl Genet, 2002, 104: 828-839[本文引用:1][JCR: 3.658]
[33]
AndersonO D, GreeneF C. The characterization and comparative analysis of high-molecular-weight glutenin genes from genomes A and B of a hexaploid bread wheat. Theor Appl Genet, 1989, 77: 689-700[本文引用:1][JCR: 3.658]
[34]
ZhongG Y, QualsetC O. Allelic diversity of high-molecular- weight glutenin protein subunits in natural populations of Dasypyrum villosum (L. ) Cand argy. Theor Appl Genet, 1993, 86: 851-858[本文引用:1][JCR: 3.658]
[35]
FengD S, ChenF G, ZhaoS Y, XiaG M. High-molecular-weight glutenin subunit genes in decaploid Agropyron elongatum. Acta Bot Sin, 2004, 46: 489-496[本文引用:1][JCR: 0.599]
[36]
封德顺, 陈凡国, 赵双宜, 夏光敏. 高冰草中一种新型高分子量谷蛋白亚基编码序列的研究. 西北植物学报, 2004, 24: 237-242FengD S, ChenF G, ZhaoS Y, XiaG M. Study on a novel HMW glutenin subunit coding region from Agropyron elongatum. Acta Bot Boreali-Occident Sin, 2004, 24: 237-242 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[37]
WangJ R, YanZ H, WeiY M, ZhengY L. A novel high- molecular-weight glutenin subunit gene Ee1. 5 from Elytrigia elongate (Host) Nevski. J Cereal Sci, 2004, 40: 289-294[本文引用:1][JCR: 2.088]
[38]
WangJ R, YanZ H, WeiY M, ZhengY L. Characterization of high-molecular-weight glutenin subunit genes from Elytrigia elongata. Plant Breed, 2006, 125: 89-95[本文引用:1][JCR: 1.175]
[39]
GuoZ F, YanZ H, WangJ R, WeiY M, ZhengY L. Characterizationof HMW prolamines and their coding sequences from Crithopsis delileana. Hereditas, 2005, 142: 56-64[本文引用:1][CJCR: 0.928]
[40]
LiuS W, ZhaoS Y, ChenF G, XiaG M. Generation of novel high quality HMW-GS genes in two introgression lines of Triticum aestivum/Agropyron elongatum. BMC Evol Biol, 2007, 7: 76[本文引用:1][JCR: 3.285]
[41]
LiuS W, GaoX, XiaG M. Characterizing HMW-GS alleles of decaploid Agropyron elongatum in relation to evolution and wheat breeding. Theor Appl Genet, 2008, 116: 325-334[本文引用:1][JCR: 3.658]
[42]
JiangQ T, WeiY M, LuZ X, PuZ E, LanX J, ZhengY L. Structural variation and evolutionary relationship of novel HMW glutenin subunits from Elymus glaucus. Hereditas, 2010, 147: 136-141[本文引用:1][CJCR: 0.928]
 , 陈其皎
, 陈其皎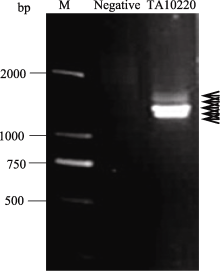









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}