关键词:siRNA; DNA甲基化; OsPCR1; 水稻; Cd积累 Comparison of Cadmium-Accumulation-Associated Genes Expression and Molecular Regulation Mechanism between Two Rice Cultivars (Oryza sativa L. subspeciesjaponica) HUANG Zhi-Xiong1,2, WANG Fei-Juan2, JIANG Han2, LI Zhi-Lan3, DING Yan-Fei2, JIANG Qiong2, TAO Yue-Liang4, ZHU Cheng1,2,* 1 State Key Laboratory of Plant Physiology and Biochemistry, College of Life Sciences, Zhejiang University, Hangzhou 310058, China
2 Zhejiang Provincial Key Laboratory of Biometrology and Inspection and Quarantine, College of Life Sciences, China Jiliang University, Hangzhou 310018, China
3Nature Science Foundation Committee of Zhejiang Province, Hangzhou 310012, China
4 College of Life and Environment Sciences, Wenzhou University, Wenzhou 325035, China
Fund: AbstractIn plants, as in other eukaryotes, endogenous small interfering RNAs (siRNAs), a class of small non-coding RNAs, and DNA methylation regulate gene expression in developmental processes and adaptating to environmental stresses, including Cd stress. Cadmium (Cd) is a non-essential heavy metal and highly toxic to plants. To investigate the regulatory role of siRNAs and DNA methylation on genes involved in heavy metals transport, we compared these genes’ expression profiles between a high Cd-accumulating rice (Oryza sativa L. subspeciesjaponica) cultivar (Xiushui 11) and a low Cd-accumulating rice cultivar (Xiushui 110). At five rice development stages investigated, the difference of these genes expression level between the two rice cultivars was not significant exceptOsPCR1, indicatingOsPCR1 may be important in Cd transport in rice. Furthermore, quantitative real time PCR (qRT-PCR) was performed to examine the expression level of a siRNA matchedOsPCR1 second exon. Results indicated that the expression level of the siRNA negatively correlated withOsPCR1 expression level at the five stages. In addition, McrBC-qRT-PCR technology was used to determine DNA methylation level, showing thatOsPCR1 expression level also negatively correlated withOsPCR1 second exon methylation level. These results of regulatory roles of siRNA and DNA methylation onOsPCR1 expression will contribute to the studies onOsPCR1 function and rice breeding for low Cd accumulation.
Keyword:siRNA; DNA methylation; OsPCR1; Rice (Oryza sativa L.); Cadmium accumulation Show Figures Show Figures
表2 5 μmol L-1 CdCl2处理24 h后, 秀水110和秀水11叶片中Cd积累相关基因的表达水平(营养生长期)(平均值± SE) Table 2 Expression level of Cd-accumulation-associated genes in leaves of Xiushui 110 and Xiushui 11 after 24 h of 5 μmol L-1CdCl2treatment (vegetative stage) (mean ± SE)
位点 Locus
秀水110 Xiushui 110
秀水11 Xiushui 11
位点 Locus
秀水110 Xiushui 110
秀水11 Xiushui 11
OsPCR1
0.81±0.12
1.71±0.08***
OsLCT1
1.49±0.30
1.05±0.11
OsPCR6
1.12±0.30
1.19±0.24
OsDCT1
1.30±0.44
1.00±0.16
OsPCR9
0.50±0.27
0.61±0.14
OsPCs
1.45±0.25
1.47±0.25
OsLCD
1.37±0.50
0.90±0.03
OsMT1
0.98±0.17
0.58±0.06
***表示秀水110和秀水11之间具有极显著的差异( P<0.001, t检验); 数据以 OsUBQ5为内参进行标准化定量, 以各自秀水110的样本为参照, n = 3~8。 *** represents a significant difference between Xiushui11 and Xiushui110 ( P<0.001, t-test); The data were normalized with OsUBQ5as reference and relative to the Xiushui110 samples respectively, n = 3 to n = 8.
表2 5 μmol L-1 CdCl2处理24 h后, 秀水110和秀水11叶片中Cd积累相关基因的表达水平(营养生长期)(平均值± SE) Table 2 Expression level of Cd-accumulation-associated genes in leaves of Xiushui 110 and Xiushui 11 after 24 h of 5 μmol L-1CdCl2treatment (vegetative stage) (mean ± SE)
表3 Table 3 表3(Table 3)
表3 5 μmol L-1 CdCl2处理60 d后秀水110和秀水11叶片中Cd积累相关基因的表达水平(抽穗期)(平均值± SE) Table 3 Expression level of Cd-accumulation-associated genes in leaves of Xiushui 110 and Xiushui 11 after 60 days of 5 μmol L-1 CdCl2treatment (pre-heading stage) (mean ± SE)
位点 Locus
秀水110 Xiushui 110
秀水11 Xiushui 11
OsPCR1
1.60±0.23
0.75±0.09**
OsLCT1
2.45±1.14
0.49±0.04
OsDCT1
1.52±0.62
0.54±0.08
**表示秀水110和秀水11之间具有显著的差异( P<0.01, t检验)。数据以 OsUBQ5为内参进行标准化定量, 以各自秀水110的样本为参照, n = 3~8。 ** represents a significant difference between Xiushui 11 and Xiushui 110 ( P<0.01, t-test). The data were normalized with OsUBQ5 as reference and relative to the Xiushui 110 samples respectively, n = 3 to n = 8.
表3 5 μmol L-1 CdCl2处理60 d后秀水110和秀水11叶片中Cd积累相关基因的表达水平(抽穗期)(平均值± SE) Table 3 Expression level of Cd-accumulation-associated genes in leaves of Xiushui 110 and Xiushui 11 after 60 days of 5 μmol L-1 CdCl2treatment (pre-heading stage) (mean ± SE)
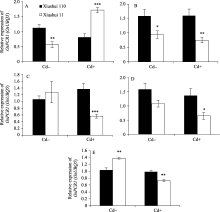
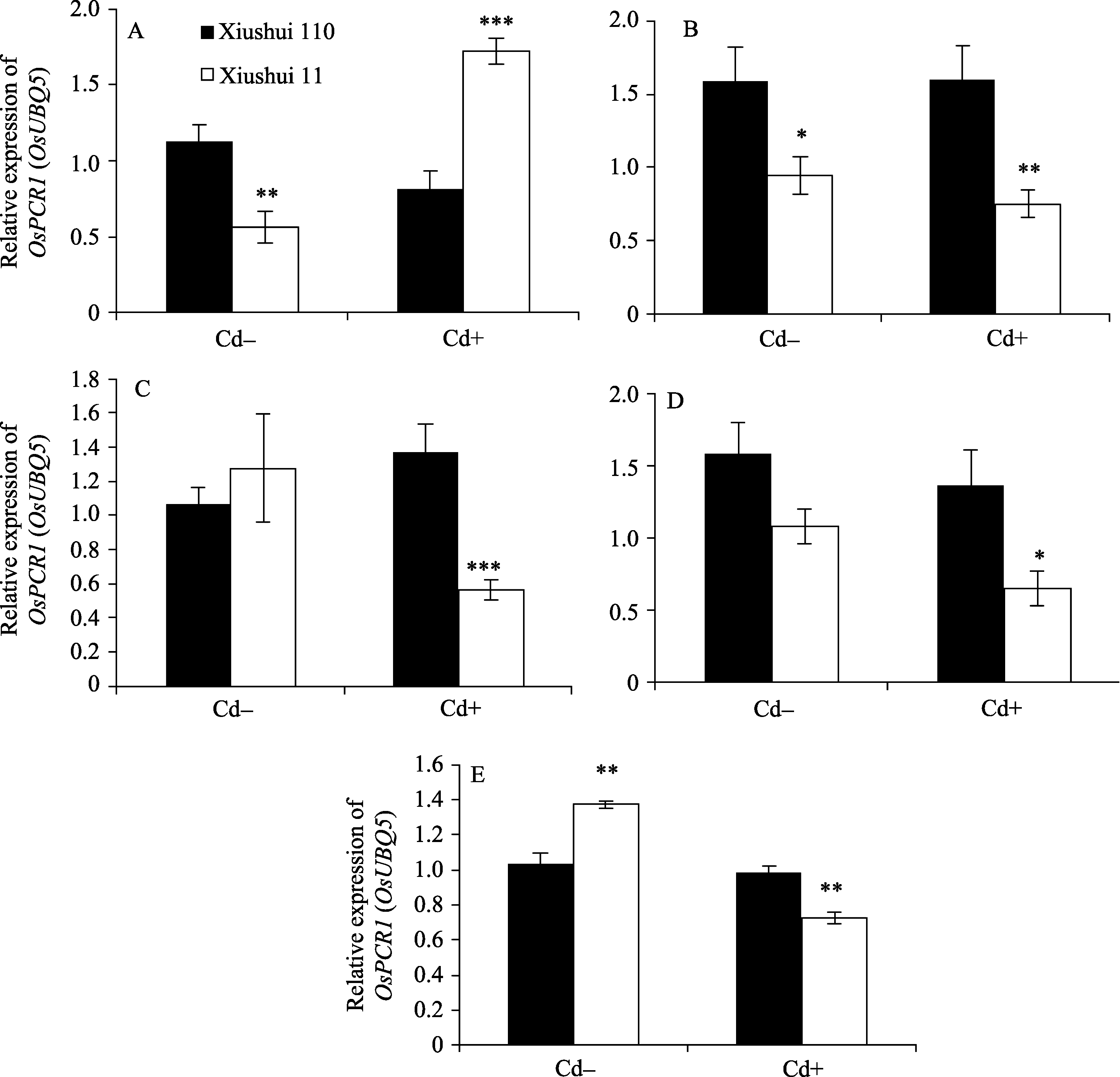
图1 不同生长发育期的秀水110和秀水11叶片中 OsPCR1基因的表达水平提取(A)营养生长期(5 μmol L-1 Cd处理1 d)、(B)抽穗前期(5 μmol L-1 Cd处理35 d)、(C)抽穗期(5 μmol L-1 Cd处理60 d)、(D)成熟期(5 μmol L-1 Cd处理105 d)和(E)完全成熟期(5 μmol L-1 Cd处理130 d)水稻品种倒数第2叶和第3叶总RNA, 没有Cd处理的样本为对照组。利用qRT-PCR检测 OsPCR1基因表达水平。数据以 OsUBQ5为内参进行标准化定量, 以各自秀水110的样本为参照, 以平均值±标准误表示(mean ± SE) ( n = 3~8)。星号表示秀水110和秀水11之间具有显著差异(* P<0.05;** P<0.01;*** P<0.001; t检验)。 Cd+: 5 μmol L-1 CdCl2处理; Cd-: 对照。Fig. 1 OsPCR1 expression in leaves of Xiushui 110 and Xiushui 11 at five developmental stages of riceRNA was isolated from the penultimate and antepenultimate leaves of the two cultivars at different development stages, including (A) vegetative stage, (B) pre-heading stage, (C) heading stage, (D) maturity stage, and (E) full-ripe stage. Cultivars at these stages were treated with 5 μmol L-1 CdCl2 for 1 d, 35 d, 60 d, 105 d, and 130 d, respectively, and cultivars without CdCl2 treatment were used as control. OsPCR1 expression was determined by qRT-PCR. Relative OsPCR1 mRNA expression was normalized with OsUBQ5 as reference and relative to the Xiushui 110 samples, respectively. The data are presented as mean ± SE ( n = 3 to n = 8). Asterisks represent a significant difference between Xiushui 11 and Xiushui 110 (* P<0.05;** P<0.01;*** P<0.001; t-test). Cd+: with 5 μmol L-1 CdCl2 treatment; Cd-: control.
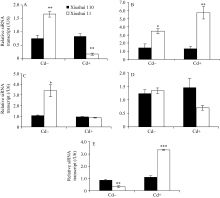
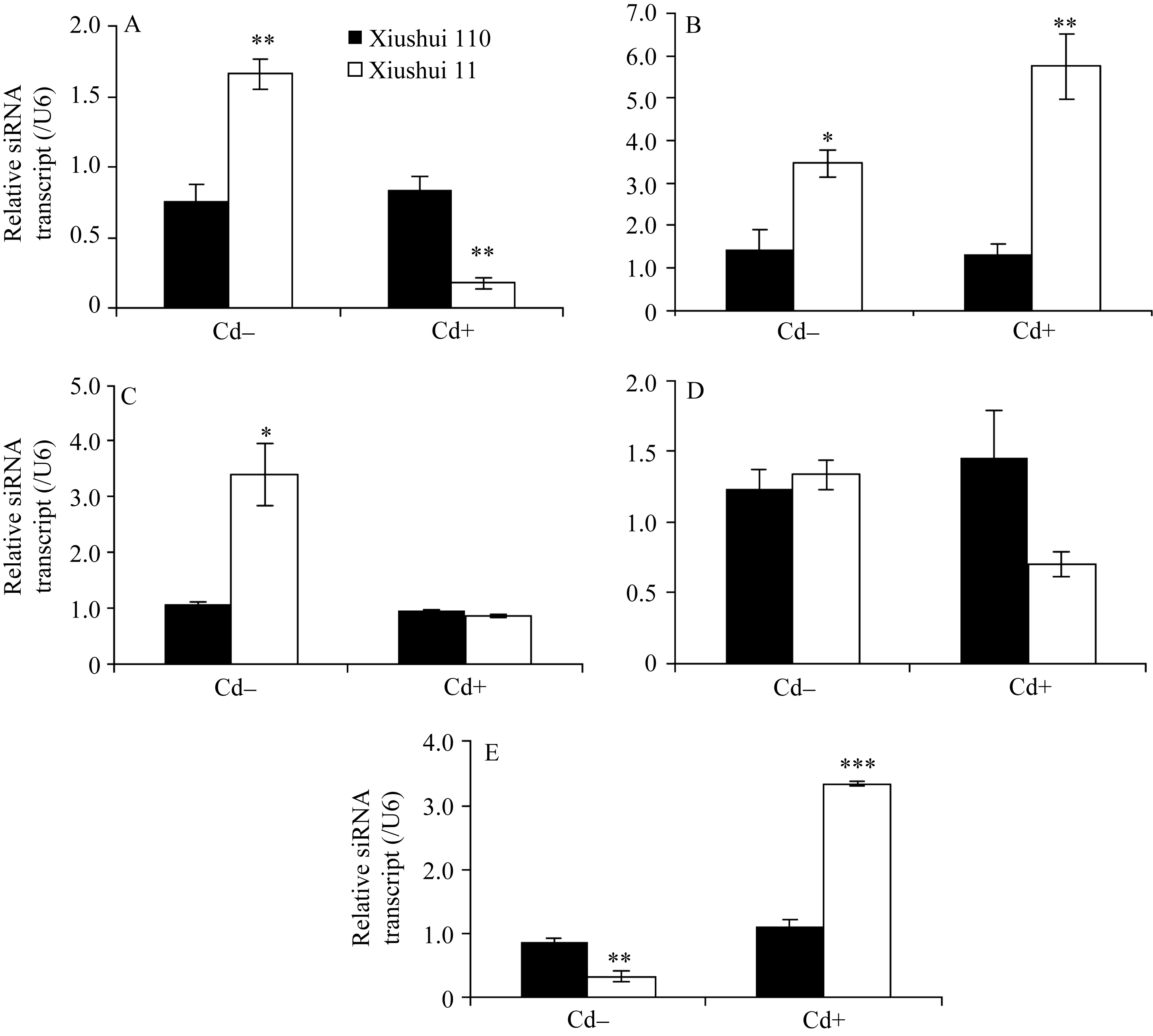
图2 不同生长发育期的秀水110和秀水11叶片中siRNA基因的表达水平利用qRT-PCR技术检测水稻品种倒数第2叶和第3叶总siRNA表达水平。数据以U6核小RNA为内参进行标准化定量, 以各自秀水110的样本为参照(定义为1), 以平均值±标准误表示(mean ± SE) ( n = 3~4)。A~E对应时期, 标注及缩写同图1。Fig. 2 siRNA expression in leaves of Xiushui 110 and Xiushui 11 at five developmental stages of riceqRT-PCR was performed to detect the expression of the siRNA in the penultimate and antepenultimate leaves of the two cultivars. Relative expression of siRNA in the leaves of Xiushui 11 was normalized to that in the leaves of Xiushui 110, which is defined as 1. U6 small nuclear RNA was used for normalization. The data are presented as mean ± SE. ( n = 3 and n = 4). The stages in individual figure, symbols and abbreviation are the same as those given in Fig. 1.
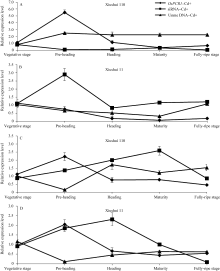
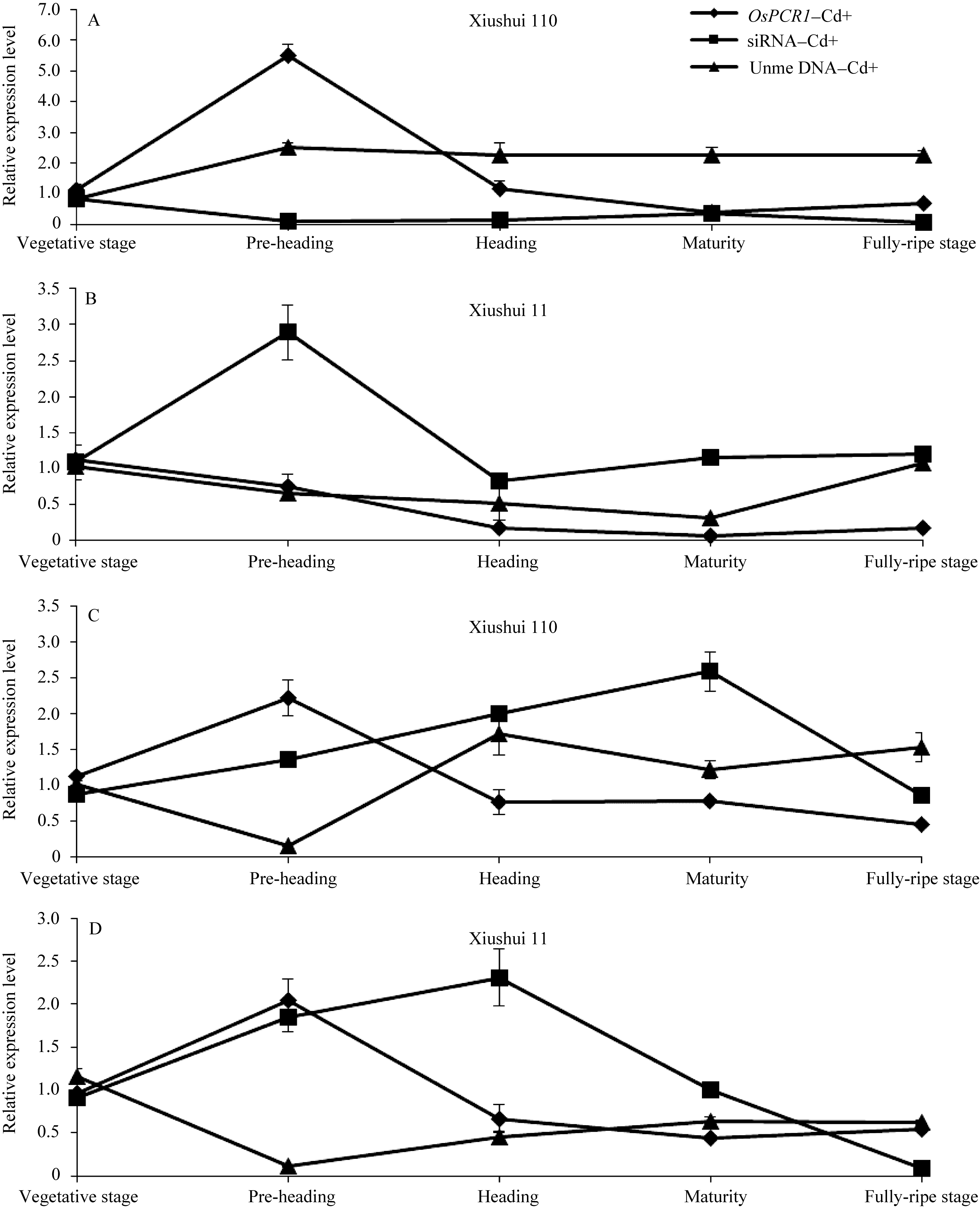
图3 不同生长发育期的秀水110和秀水11叶片中 OsPCR1基因、siRNA的表达水平和DNA甲基化水平提取5个不同生长发育期秀水110和秀水11倒数第2和第3叶基因组DNA和总RNA。利用McrBC-qRT-PCR检测 OsPCR1外显子2甲基化水平, qRT-PCR检测 OsPCR1基因表达水平和siRNA表达水平。相对 OsPCR1外显子2甲基化水平的数据校准到等量未经酶切的基因组DNA。 OsPCR1基因的表达水平校准到 OsUBQ5。siRNA的表达水平校准到U6小核仁RNA。所有数据以各自营养生长期的样本为参照, 以平均值±标准误表示(mean ± SE) ( n=3~8)。 OsPCR1: OsPCR1相对表达水平; siRNA: siRNA相对表达水平; Unme: OsPCR1外显子2相对未甲基化水平。标注及缩写同图1。Fig. 3 OsPCR1 expression level, siRNA expression level, and OsPCR1 exon 2 methylation level in leaves of (A) Xiushui 110 and (B) Xiushui 11 at five developmental stages of riceGenomic DNA and total RNA were isolated from leaves previously described in Fig. 1 legend. OsPCR1 exon 2 methylation, OsPCR1 and siRNA expressions were determined by the methods of McrBC-qRT-PCR and qRT-PCR, respectively. Relative unmethylated OsPCR1 exon 2 level, OsPCR1 mRNA and siRNA expressions were normalized to equal amounts of undigested DNA samples, OsUBQ5 and U6 small nuclear RNA, respectively and relative to the vegetative samples. The data are presented as mean ± SE ( n = 3 to n = 8). OsPCR1: relative OsPCR1expression level; siRNA: relative siRNA level; Unme: relative unmethylated OsPCR1 exon 2 level. Symbols and abbreviation are the same as those given in Fig. 1.
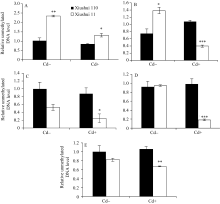
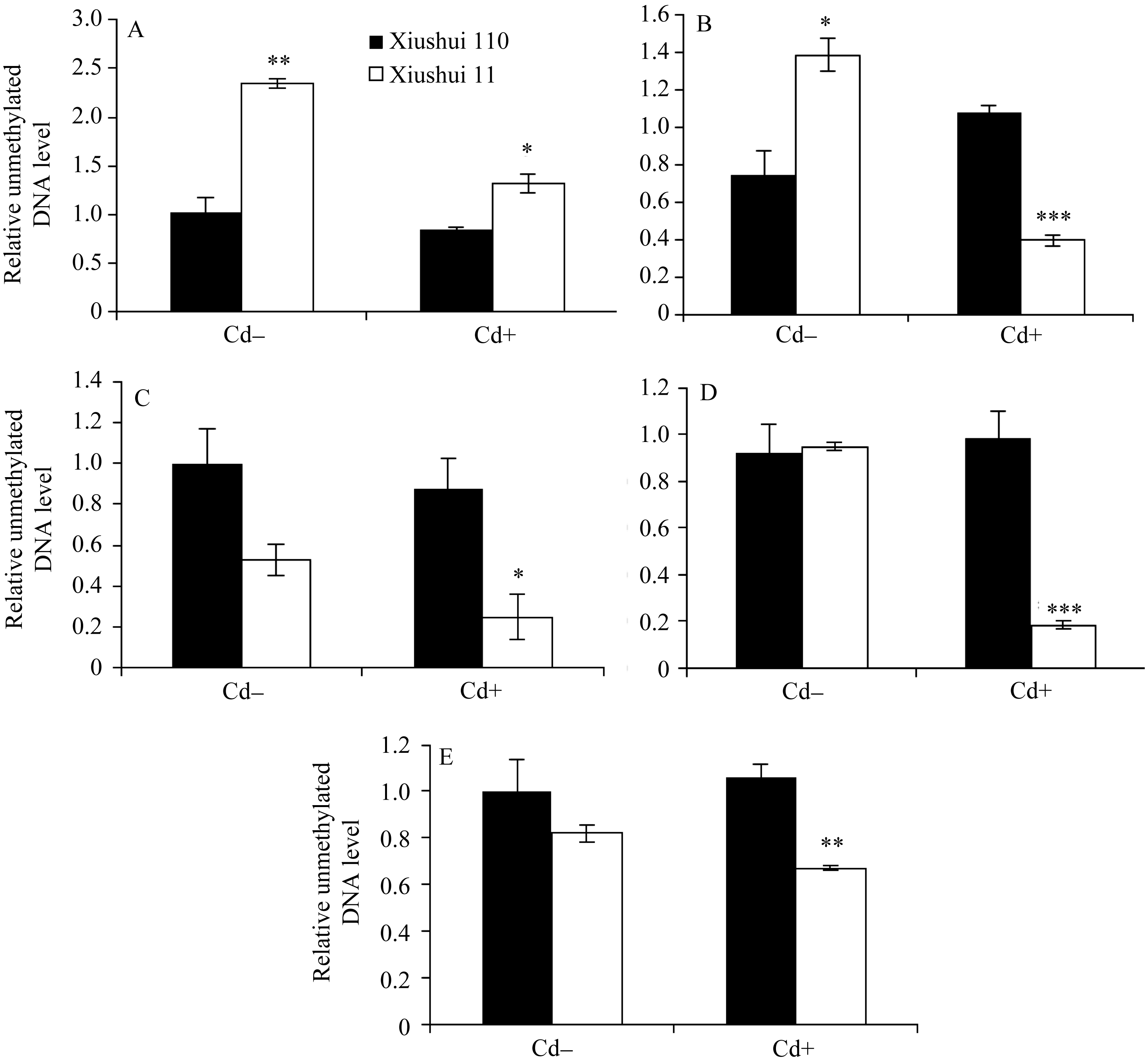
图4 不同生长发育期的秀水110和秀水11叶片中 OsPCR1基因外显子2甲基化水平提取5个不同生长发育期秀水110和秀水11倒数第2和第3叶基因组DNA。利用特异性的引物和McrBC-qRT-PCR技术检测 OsPCR1基因外显子2甲基化水平。数据校准到等量未经酶切的DNA样本, 以各自秀水110的样本为参照(定义为1), 以平均值±标准误表示(mean ± SE) ( n = 3~4)。A~E对应时期, 标注及缩写同图1。Fig. 4 OsPCR1exon 2 methylation in leaves of Xiushui 110 and Xiushui 11 at five developmental stages of riceGenomic DNA was isolated from leaves described in Fig. 1 legend. OsPCR1exon 2 methylation was determined by the methods of McrBC-qRT-PCR with specific primers. The data were normalized to equal amount of undigested DNA samples and relative to Xiushui 110 samples, respectively. The data are presented as means ± SE ( n = 3 and n = 4). The stages in individual figure, symbols and abbreviation are the same as those given in Fig. 1.
UraguchiS, KamiyaT, SakamotoT, KssaiK, SatoY, NagamuraY, YoshidaA, KyozukaJ, IshikawaS, FujiwaraT. Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains. Proc Natl Acad Sci USA, 2011, 108: 20959-20964[本文引用:4][JCR: 9.737]
[2]
UenoD, KoyamaE, YamajiN, MaJ F. Physiological, genetic, and molecular characterization of a high-Cd-accumulating rice cultivars, Jarjan. J Exp Bot, 2011, 62: 2262-2272[本文引用:1][JCR: 5.242]
[3]
UraguchiS, MoriS, KuramataM, KawasakiA, AraoT, IshikawaS. Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. J Exp Bot, 2009, 60: 2677-2688[本文引用:1][JCR: 5.242]
[4]
TakahashiR, IshimaruY, SenouraT, ShimoH, IshikawaS, AraoT, NakanishiH, NishizawaN K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J Exp Bot, 2011, 62: 4843-4850[本文引用:1][JCR: 5.242]
[5]
KuramataM, MasuyaS, TakahashiY, KitagawaE, InoueC, IshikawaS, YoussefianS, KusanoT. Novel cysteine-rich peptides from Digitaria ciliaris and Oryza sativa enhance tolerance to cadmium by limiting its cellular accumulation. Plant Cell Physiol, 2009, 50: 106-117[本文引用:3][JCR: 4.134]
[6]
ShenG M, ZhuC, DuQ Z. Genome-wide identification of PHYTOCHELATIN and PHYTOCH_SYNTH domain-containing phytochelatin family from rice. Electronic J Biol, 2010, 6: 73-79[本文引用:2]
[7]
ShimoH, IshimaruY, AnG, YamakawaT, NakanishiH, NishizawaN K. Low cadmium (LCD), a novel gene related to cadmium tolerance and accumulation in rice. J Exp Bot, 2011, 62: 5727-5734[本文引用:3][JCR: 5.242]
[8]
SongW Y, ChoiK S, AlexisD A, MartinoiaE, LeeY. Brassica juncea plant cadmium resistance 1 protein (BjPCR1) facilitates the radial transport of calcium in the root. Proc Natl Acad Sci USA, 2010, 108: 19808-19813[本文引用:3][JCR: 9.737]
[9]
SongW Y, ChoiK S, KimD Y, GeislerM, ParkJ, VincenzettiV, SchellenbergM, KimS H, LimY P, NohE W, LeeY, MartinoiaE. Arabidopsis PCR2 is a zinc exporter involved in both zinc extrusion and long-distance zinc transport. Plant Cell, 2010, 22: 2237-2252[本文引用:5][JCR: 9.251]
[10]
CobbettC, GoldsbroughP. Phytochelatins and metallothioneins: roles in heavy metal detoxification and homeostasis. Annu Rev Plant Biol, 2002, 53: 159-182[本文引用:1][JCR: 25.962]
[11]
SongW Y, HörtensteinerS, TomiokaR, LeeY, MartinoiaE. Common functions or only phylogenetically related? The large family of PLAC8 motif-containing/PCR genes. Mol Cells, 2011, 31: 1-7[本文引用:1][JCR: 2.21]
[12]
SongW Y, MartinoiaE, LeeJ, KimD, KimD Y, VogtE, ShimD, ChoiK S, HwangI, LeeY. A novel family of cys-rich membrane proteins mediates cadmium resistance in Arabidopsis. Plant Physiol, 2004, 135: 1027-1039[本文引用:3][JCR: 6.555]
[13]
BorsaniO, ZhuJ, VersluesP E, SunkarR, ZhuJ K. Endogenous siRNAs derived from a pair of natural cis-antisense transcripts regulate salt tolerance in Arabidopsis. Cell, 2005, 123: 1279-1291[本文引用:2][JCR: 31.957]
[14]
CarthewR W, SontheimerE J. Origins and mechanisms of miRNA and siRNAs. Cell, 2009, 136: 642-655[本文引用:2][JCR: 31.957]
[15]
MoldovanD, SpriggsA, YangJ, PogsonB J, DennisE S, WilsonI W. Hypoxia-responsive microRNAs and trans-acting small interfering RNAs in Arabidopsis. J Exp Bot, 2010, 61: 165-177[本文引用:2][JCR: 5.242]
[16]
YanY, ZhangY, SunZ, FuY, ChenX, FangR. Small RNAs from MITE-derived stem-loop precursors regulate abscisic acid signaling and abiotic stress responses in rice. Plant J, 2011, 65: 820-828[本文引用:2][JCR: 6.582]
[17]
YaoY, NiZ, PengH, SunF, XinM, SunkarR, ZhuJ K, SunQ. Non-coding small RNAs responsive to abiotic stress in wheat (Triticum aestivum L. ). Funct Integr Genomic, 2010, 10: 187-190[本文引用:2][JCR: 3.292]
[18]
KimV N, HanJ, SiomiM C. Biogenesis of small RNAs in animals. Nat Rev Mol Cell Biol, 2009, 10: 126-139[本文引用:3]
[19]
HannonG J. RNA interference. Nature, 2002, 418: 244-251[本文引用:1][JCR: 38.597]
[20]
SongJ J, SmithS K, HannonG J, Joshua-TorL. Crystal structure of Argonaute and its implications for RISC slicer activity. Science, 2004, 305: 1434-1437[本文引用:1]
[21]
LawJ A, JacobsenS E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat Rev Genet, 201011: 204-220[本文引用:3]
[22]
Chan S W L, HendersonI R, JacobsenS E. Gardening the genome: DNA methylation in Arabidopsis thaliana. Nat Rev Genet, 2005, 6: 351-360[本文引用:3][JCR: 41.063]
[23]
BoykoA, BlevinsT, YaoY, GolubovA, BilichakA, IlnytskyyY, Holland erJ. Transgenerational adaptation of Arabidopsis to stress requires DNA methylation and the function of dicer-like proteins. PLoS One, 2010, 5: e9514[本文引用:2][JCR: 3.73]
[24]
ChoiC S, SanoH. Abiotic-stress induces demethylation and transcriptional activation of a gene encoding a glycerophosphodiesterase-like protein in tobacco plants. Mol Genet Genomics, 2007, 277: 589-600[本文引用:2][JCR: 2.881]
[25]
GrecoM, ChiappettaA, BrunoL, BitontiM B. In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning. J Exp Bot, 2012, 63: 695-709[本文引用:2][JCR: 5.242]
[26]
Verhoeven K J F, JansenJ J, DijkP J, BiereA. Stress-induced DNA methylation changes and their heritability in asexual dand elions. New Phytol, 2010, 185: 1108-1118[本文引用:2][JCR: 6.736]
[27]
RajS, BrautigenK, HamanishiE T, WilkinsO, ThomasB R, SchroederW, MansfieldS D, PlantA L, CampbellM M. Clone history shapes populous drought response. Proc Natl Acad Sci USA, 2011, 108: 12521-12526[本文引用:2][JCR: 9.737]
[28]
BallM P, LiJ B, GaoY, LeeJ H, LeProustE M, ParkI H, XieB, DaleyG Q, ChurchG M. Targeted and genome-scale strategies reveal gene-body methylation signatures in human cells. Nat Biotechnol, 2009, 27: 361-368[本文引用:1][JCR: 32.438]
[29]
HellmanA, ChessA. Gene Body-specific methylation on the active X chromosome. Science, 2007, 315: 1141-1143[本文引用:2]
[30]
ListerR, PelizzolaM, DowenR H, HawkinsR D, HonG, Tonti-FilippiniJ, NeryJ R, LeeL, YeZ, NgoQ M, EdsallL, Antosiewicz-BourgetJ, StewartR, RuottiV, MillarA H, ThomsonJ A, RenB, EckerJ R. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature, 2009, 462: 315-322[本文引用:1][JCR: 38.597]
[31]
KawanabeT, FujimotoR, SasakiT, TaylorJ M, DennisE S. A comparison of transcriptome and epigenetic status between closely related species in the genus Arabidopsis. Gene, 2012, 506: 301-309[本文引用:1][JCR: 2.196]
[32]
ZilbermanD, GehringM, TranR K, BallingerT, HenikoffS. Genome-wide analysis of Arabidopsis thaliana DNA methylation uncovers an interdependence between methylation and transcription. Nat Genet, 2007, 39: 61-69[本文引用:2][JCR: 35.209]
[33]
SuzukiM M, BirdA. DNA methylation land scapes: provocative insights from epigenomics. Nat Rev Genet, 2008, 9: 465-476[本文引用:1][JCR: 41.063]
何俊瑜, 任艳芳, 朱诚, 蒋德安. 镉胁迫对不同水稻品种种子萌发、幼苗生长和淀粉酶活性的影响. 中国水稻科学, 2008, 22: 399-404HeJ Y, RenY F, ZhuC, JiangD A. Effects of cadmium stress on seed germination, seedling growth, and amylase activities in rice. Chin J Rice Sci, 2008, 22: 399-404 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[36]
DingY, ChenZ, ZhuC. Microarray-based analysis of cadmium-responsive microRNAs in rice (Oryza sativa). J Exp Bot, 2011, 62: 3563-3573[本文引用:2][JCR: 5.242]
[37]
PfafflM W. A new mathematical model for relative quantification in real-time RT-PCR. Nucl Acids Res, 2001, 29: e45[本文引用:3]
[38]
SerraI A, ProcacciniG, IntrieriM C, MigliaccioM, MazzucaS, InnocentiA M. Comparison of ISSR and SSR markers for analysis of genetic diversity in the seagrass Posidonia oceanica. Marine Ecol Prog Ser, 2007, 338: 71-79[本文引用:1][JCR: 2.546]
[39]
TeixeiraF K, HerediaF, SarazinA, RoudierF, BoccaraM, CiaudoC, CruaudC, PoulainJ, BerdascoM, FragaM F, VoinnetO, WinckerP, EstellerM, ColotV. A role for RNAi in the selective correction of DNA methylation defects. Science, 2009, 323: 1600-1604[本文引用:1]
[40]
JohnsonC, BowmanL, AdaiA T, VanceV, SundaresanV. CSRDB: a small RNA integrated database and browser resource for cereals. Nucl Acids Res, 2007, 35: D829-D833[本文引用:1]
[41]
Brand eisM, ArielM, CedarH. Dynamics of DNA methylation during development. Bioessays, 1993, 15: 709-713[本文引用:1][JCR: 5.423]
[42]
赵嵘, 胡丽玲, 孔繁强, 左爱军. PXR基因外显子3甲基化与肠癌细胞对5氟尿嘧啶的耐药性相关. 中国生物化学与分子生物学报, 2013, 29: 63-69ZhaoR, HuL L, KongF Q, ZuoA J. Association between pregnane X receptor gene exon3 methylation with 5-fluorouracil resistance of the colon cancer cells. Chin J Biochem Mol Biol, 2013, 29: 63-69 (in Chinese with English abstract)[本文引用:1][CJCR: 0.623]
[43]
HohnT, CorstenS, RiekeS, MullerM, RothnieH. Methylation of coding region alone inhibits gene expression in plant protoplasts. Proc Natl Acad Sci USA, 1996, 93: 8334-8339[本文引用:1][JCR: 9.737]
[44]
RountreeM R, SelkerE U. DNA methylation inhibits elongation but not initiation of transcription in Neurospora crassa. Genes Dev, 1997, 11: 2383-2395[本文引用:1][JCR: 12.444]
[45]
XiaoZ, WangC, MoD, LiJ, ChenY, ZhangZ, CongP. Promoter CpG methylation status in porcine Lyn is associated with its expression levels. Gene, 2012, 511: 73-78[本文引用:1][JCR: 2.196]
[46]
ForetS, KucharskiR, PellegriniM, FengS, JacobsenS E, RobinsonG E, MaleszkaR. DNA methylation dynamics, metabolic fluxes, genesplicing, and alternative phenotypes in honey bees. Proc Natl Acad Sci USA, 2012, 109: 4968-4973[本文引用:1][JCR: 9.737]
[47]
ShuklaS, KavakE, GregoryM, ImashimizuM, ShutinoskiB, KashlevM, OberdoerfferP, Sand bergR, OberdoerfferS. CTCF-promoted RNA polymerase II pausing links DNA methylation to splicing. Nature, 2011, 479: 74-79[本文引用:1][JCR: 38.597]


{kind=link}
{kind=link}
{kind=link}
{kind=link}