关键词:大豆; 育成品种; SSR; 遗传多样性; 特异性 Genetic Similarity and Specificity of Ten Important Soybean Cultivar Families Released in China XIONG Dong-Jin1,2,**, WANG Wu-Bin1,**, ZHAO Tuan-Jie1, GAI Jun-Yi1,* 1Soybean Research Institute of Nanjing Agricultural University / National Center for Soybean Improvement / National Key Laboratory for Crop Genetics and Germplasm Enhancement, Nanjing 210095, China
2 School of Life Science and food Engineering, Nanchang University, Nanchang 330031, China
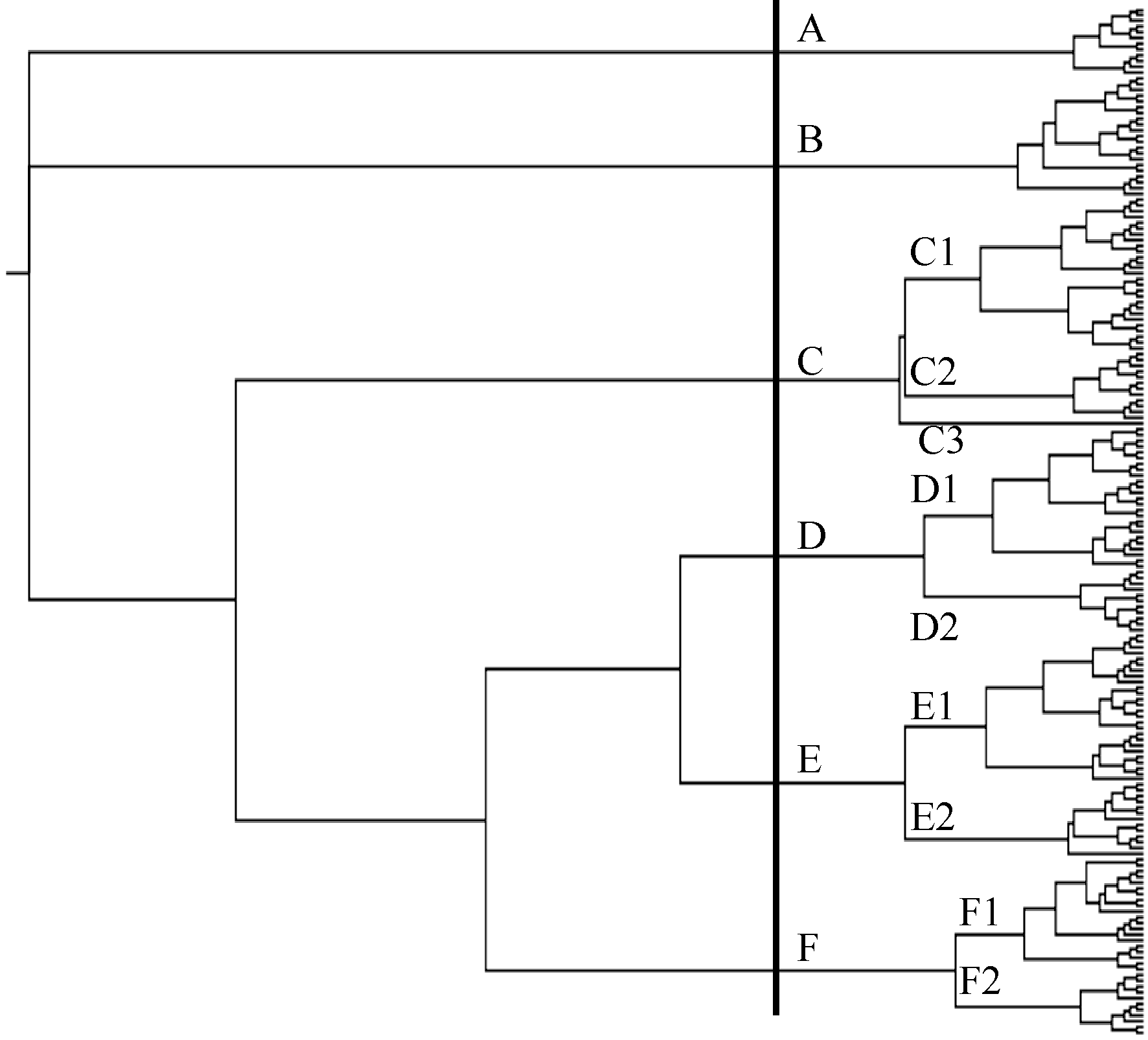
Fund: AbstractAnalysis of the affinity relationship and genetic similarity among cultivars is important for crop genetic improvement. In this study, a total of 161 SSRs covering the entire soybean genome were analyzed for the genetic diversity, similarity and specificity of 10 important families composed of 179 cultivars using PowerMarker Ver. 3.25. The results showed that there were totally 1697 alleles, averaging 10.5 per locus, ranging from 5 to 24 with average polymorphism information content of 0.832, ranging from 0.545 to 0.943 in the population. According to the SSR cluster analysis, the 179 cultivars were clustered into six groups, eleven subgroups, with a tendency that the cultivars in a family tended to be grouped into a same cluster. There existed significant correlation (r= 0.67) between coefficient of parentage (CP) and genetic similarity coefficient (GSC) of the population. The CP and GSC values of A295, A133, A122, and A231 families were relatively low, which means that the genetic bases of the four families were relatively broad. In contrast, the CP and GSC values of A291, A201, A084, and A002 families were relatively high, indicating their genetic bases were relatively narrow due to more cultivars obtained from pure line selection. The CP and GSC values between the A019 family from Northeast China and other nine families were the lowest among all pairwise combinations of families. The genetic specificity analysis showed that there existed much more complementary alleles, specifically existent and specifically deficient alleles in A019 in comparison with in other families, indicating the former is distant from the latter. On the other hand, the families in Eco-region II and III, contained fewer complementary alleles, specifically existent and specifically deficient alleles, which might be due to some frequent germplasm exchange between the neighboring eco-regions. For example, there were no specifically existent alleles in A002, A231, A122 and no specifically deficient alleles in A084, A201, A034, and A231. The present results are of significance in broadening the genetic basis of soybean cultivar.
Keyword:Soybean; Released cultivar; SSR; Genetic diversity; Specificity Show Figures Show Figures
I: 北方一熟春豆生态区; II: 黄淮海二熟春夏豆生态区; III: 长江中下游二熟春夏豆生态区; IV: 中南多熟春夏秋豆生态区; V: 西南高原二熟春夏豆生态区; 华南热带多熟多播季生态区(VI)的大豆未包括在本研究中。 I: Northern single cropping, spring planting eco-region; II: Huanghuaihai double cropping, spring and summer planting eco-region; III: Middle and lower Changjiang valley double cropping, spring and summer planting eco-region; IV: Central south multiple cropping, spring, summer and autumn planting eco-region; V: Southwest plateau double cropping, spring and summer planting eco-region; cultivars from South China tropical multiple cropping, all season planting eco-region (VI) is excluded in the present study.
表5 主要家族间亲本系数和遗传相似系数的比较 Table 5 Comparison of parentage coefficient and genetic similarity coefficient among the families
家族Family
A019
A295
A133
A122
A034
A231
A002
A084
A201
A291
A019
0.14
0.14
0.14
0.14
0.14
0.13
0.14
0.13
0.13
A295
0
0.17
0.18
0.17
0.17
0.16
0.16
0.16
0.17
A133
0
0.05
0.17
0.16
0.16
0.15
0.16
0.16
0.16
A122
0
0.06
0.04
0.17
0.17
0.16
0.16
0.15
0.17
A034
0
0.06
0.04
0.04
0.17
0.16
0.16
0.16
0.17
A231
0
0.04
0.04
0.04
0.06
0.16
0.16
0.16
0.16
A002
0.01
0.03
0.03
0.03
0.06
0.05
0.18
0.14
0.15
A084
0.01
0.03
0.02
0.03
0.03
0.03
0.11
0.16
0.16
A201
0
0.01
0.02
0.01
0.02
0.02
0.01
0.02
0.24
A291
0
0.01
0.01
0
0.02
0.02
0.01
0.03
0.09
对角线上方为遗传相似系数, 对角线下方为亲本系数。 Numbers above the diagonal are genetic similarity coefficient (GSC) values; those below the diagonal are coefficient of parentage (CP) values.
表5 主要家族间亲本系数和遗传相似系数的比较 Table 5 Comparison of parentage coefficient and genetic similarity coefficient among the families
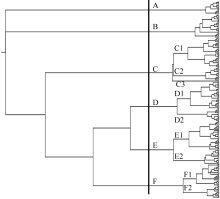
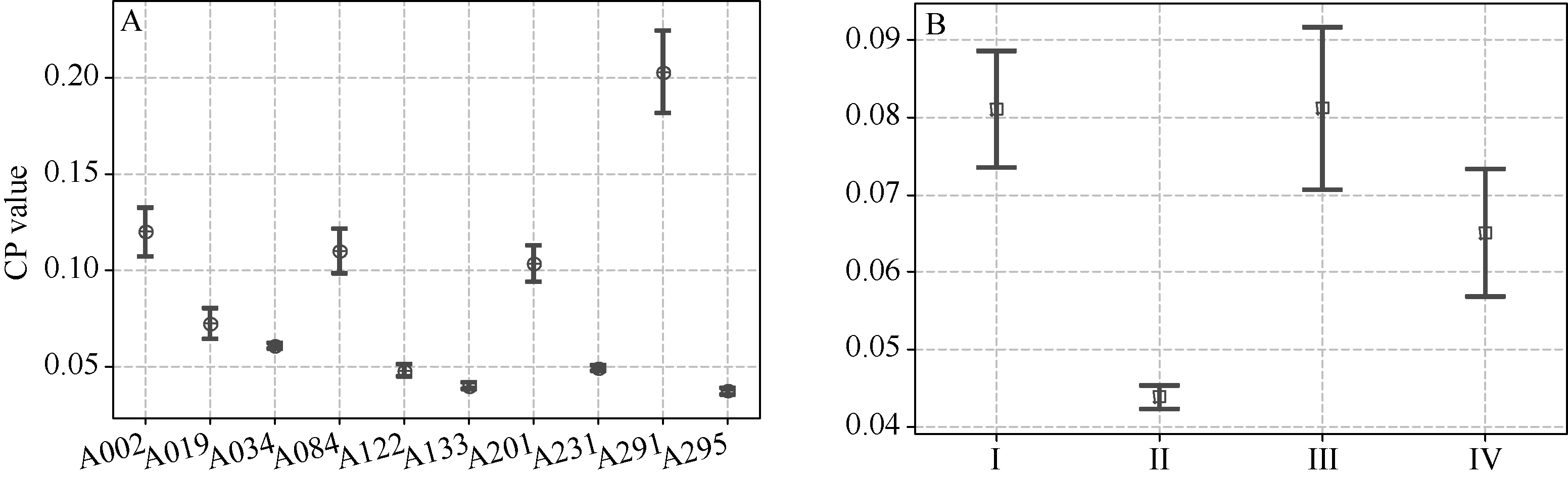
图2 成对品种间亲本系数频数分布图A: 不同家族; B: 不同生态区。Fig. 2 Frequency distribution of CP in pairwised soybean cultivarsA: frequency distribution in different families; B: frequency distribution in different eco-regions.
表7 不同家族间互补等位变异数 Table 7 Number of complementary alleles among different families
家族Family
A019
A295
A133
A122
A034
A231
A002
A084
A201
A291
A019
102
48
201
35
33
182
166
363
433
A295
604(706)
20
242
21
23
298
309
538
670
A133
642(690)
112(132)
336
43
46
339
335
593
733
A122
468(669)
7(249)
9(345)
9
11
221
211
437
522
A034
668(703)
152(173)
82(125)
375(384)
26
350
367
638
770
A231
656(689)
144(167)
75(121)
367(378)
16(42)
344
353
632
766
A002
483(665)
97(395)
46(385)
255(476)
18(368)
22(366)
131
467
573
A084
467(633)
187(496)
42(377)
245(456)
35(402)
31(384)
131(262)
459
547
A201
380(743)
53(591)
16(609)
187(624)
22(660)
26(658)
183(650)
175(634)
256
A291
331(764)
46(716)
17(750)
133(655)
15(785)
21(787)
150(723)
124(671)
117(373)
表中所列的行家族对列家族补充的等位变异数; 括号内的数字为2个亚群间互补等位变异数。 The numbers of complementary alleles in the Table are those in the vertical lines to the horizontal lines; the numbers in parentheses are the numbers of complementary alleles between two families.
表7 不同家族间互补等位变异数 Table 7 Number of complementary alleles among different families
表8 Table 8 表8(Table 8)
表8 不同家族群体间特有等位变异和特缺等位变异 Table 8 Specifically existent and specifically deficient alleles between different ancestor families
家族 Family
特有等位变异 Specifically existent allele
特缺等位变异 Specifically deficient allele
标记数 Marker number
等位变异数 Allele number
标记数 Marker number
等位变异数 Allele number
A002
0
0
14
15
A019
10
10
103
145
A034
3
3
0
0
A084
2
2
0
0
A201
3
3
0
0
A231
0
0
0
0
A291
1
1
17
17
A295
4
4
36
37
A122
0
0
60
71
A133
3
3
0
0
表8 不同家族群体间特有等位变异和特缺等位变异 Table 8 Specifically existent and specifically deficient alleles between different ancestor families
CuiZ L, CarterT E, BurtonJ W. Genetic diversity patterns in Chinese soybean cultivars based on coefficient of parentage. Crop Sci, 2000, 40: 1780-1793[本文引用:2][JCR: 1.513]
[2]
ZhouX G, CarterT E, CuiZ L, MiyazakiS, BurtonJ W. Genetic diversity patterns in Japanese soybean cultivars based on coefficient of parentage. Crop Sci, 2002, 42: 1331-1342[本文引用:2][JCR: 1.513]
[3]
盖钧镒, 崔章林. 中国大豆育成品种的亲本分析. 南京农业大学学报, 1994, 17: 19-23GaiJ Y, CuiZ L. Ancestral analysis of soybean cultivars released in China. J Nanjing Agric Univ, 1994, 17: 19-23 (in Chinese with English abstract)[本文引用:1][CJCR: 0.916]
[4]
SnellerC H. Pedigree analysis of elite soybean lines. Crop Sci, 1994, 34: 1515-1522[本文引用:1][JCR: 1.513]
[5]
BharadwajC H, SatyavathiC T, TiwariS P. Genetic base of soybean (Glycine max) varieties released in India as revealed by coefficient of parentage. Ind J Agric Sci, 2002, 72: 467-469[本文引用:1]
[6]
Bonato A L V, CalvoE S, GeraldiI O, Arias C A A. Genetic similarity among soybean (Glycine max (L. ) Merrill) cultivars released in Brazil using AFLP markers. Genet Mol Biol, 2006, 29: 692-704[本文引用:1][JCR: 0.744]
[7]
NarvelJ M, FehrW R, ChuW S, GrantD, ShoemakerR C. Simple sequence repeat diversity among soybean plant introductions and elite genotypes. Crop Sci, 2000, 40: 1452-1458[本文引用:1][JCR: 1.513]
[8]
AbeJ, XuD H, SuzukiY, KanazawaA, ShimamotoY. Soybean germplasm pools in Asia revealed by nuclear SSRs. Theor Appl Genet, 2003, 106: 445-453[本文引用:1][JCR: 3.658]
[9]
MimuraM, CoyneC J, BambuckM W, LumpkinT A. SSR diversity of vegetable soybean [Glycine max (L. ) Merr. ]. Genet Resour Crop Evol, 2007, 54: 497-508[本文引用:1][JCR: 1.593]
[10]
WangL X, GuanR X, LiY H, LinF Y, LuanW J, LiW, MaY S, LiuZ X, ChangR Z, QiuL J. Genetic diversity of Chinese spring soybean germplasm revealed by SSR markers. Plant Breed, 2008, 127: 56-61[本文引用:1][JCR: 1.175]
[11]
UdeG N, KenworthyW J, CostaJ M, CreganP B, AlvernazJ. Genetic diversity of soybean cultivars from China, Japan, North America, and North American ancestral lines determined by amplified fragment length polymorphism. Crop Sci, 2003, 43: 1858-1867[本文引用:1][JCR: 1.513]
[12]
FuY, PetersonG W, MorrisonM J. Genetic diversity of Canadian soybean cultivars and exotic germplasm revealed by simple sequence repeat markers. Crop Sci, 2007, 47: 1947-1954[本文引用:1][JCR: 1.513]
[13]
YamanakaN, SatoH, YangZ, XuD H, CatelliL L, Arias C A A, AbdelnoorR V, NepomucenoA L. Genetic relationships between Chinese, Japanese, and Brazilian soybean gene pools revealed by simple sequence repeat (SSR) markers. Genet Mol Biol, 2007, 30: 85-88[本文引用:1][JCR: 0.744]
[14]
Priolli R H G, Mendes-JuniorC T, Sousa S M B, Sousa N E A, Contel E P B. Soybean genetic diversity in time and among breeding programs in Brazil. Pesq Agropec Bras, 2004, 39: 967-975[本文引用:1]
[15]
Priolli R H G, Mendes-JuniorC T, ArantesN E, Contel E P B. Characterization of Brazilian soybean cultivars using microsatellite markers. Genet Mol Biol, 2002, 25: 185-193[本文引用:1][JCR: 0.744]
[16]
关荣霞, 郭娟娟, 常汝镇, 邱丽娟. 国外种质对中国大豆育成品种遗传贡献的分子证据. 作物学报, 2007, 33: 1393-1398GuanR X, GuoL L, ChangL Z, QiuL J. Marker-based evidence of broadening the genetic base of Chinese soybeans by using introduced soybeans. Acta Agron Sin, 2007, 33: 1393-1398 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[17]
张军, 赵团结, 盖钧镒. 亚洲大豆栽培品种遗传多样性、特异性和群体分化研究. 中国农业科学, 2008, 41: 3511-3520ZhangJ, ZhaoT J, GaiJ Y. Genetic diversity, specificity and population differentiation of soybean cultivars in Asia. Sci Agric. Sin, 2008, 41: 3511-3520 (in Chinese with English abstract)[本文引用:4]
[18]
MaccaferriM, SanguinetiM C, XieC, SmithJ S, TuberosaR. Relationships among durum wheat accessions: II. A comparison of molecular and pedigree information. Genome, 2007, 50: 385-399[本文引用:1][JCR: 1.668]
[19]
VanBecelaere G, LubbersE L, PatersonA H, CheeP W. Pedigree- vs. DNA marker-based genetic similarity estimates in cotton. Crop Sci, 2005, 45: 2281-2287[本文引用:1][JCR: 1.513]
[20]
张军, 赵团结, 盖钧镒. 我国黄淮和南方主要大豆育成品种家族产量和品质优异等位变异在系谱中遗传的研究. 作物学报, 2009, 35: 191-201ZhangJ, ZhaoT J, GaiJ Y. Inheritance of elite alleles of yield and quality traits in the pedigrees of major cultivar families released in Huanghuai Valleys and Southern China. Acta Agron Sin, 2009, 35: 191-201 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[21]
熊冬金, 赵团结, 盖钧镒. 中国大豆育成品种亲本分析. 中国农业科学, 2008, 41: 2589-2598XiongD J, ZhaoT J, GaiJ Y. Parental analysis of soybean cultivars released in China. Sci AgricSin, 2008, 41: 2589-2598 (in Chinese with English abstract)[本文引用:1][JCR: 1.44]
[22]
熊冬金, 赵团结, 盖钧镒. 1923—2005年中国大豆育成品种的核心祖先亲本分析. 大豆科学, 2007, 26: 2589-2598XiongD J, ZhaoT J, GaiJ Y. The core ancestors of soybean cultivars released during 1923-2005 in China. Soybean Sci, 2007, 26: 2589-2598 (in Chinese with English abstract)[本文引用:1]
[23]
AljanabiS M, ForgetL, DookunA. An improved and rapid protocol for the isolation of polysaccharide- and polyphenol-free sugarcane DNA. Plant Mol Biol Rep, 1999, 17: 181[本文引用:1][JCR: 5.319]
[24]
SongQ J, MarekL F, ShoemakerR C, LarkK G, ConcibidoV C, DelannayX, SpechtJ E, CreganP B. A new integrated genetic linkage map of the soybean. Theor Appl Genet, 2004, 109: 122-128[本文引用:1][JCR: 3.658]
[25]
CoxT S, KiangY T, GormanM B, RodgersD M. Relationship between coefficient of parentage and genetic similarity indices in the soybean. Crop Sci, 1985, 25: 529-532[本文引用:1][JCR: 1.513]
[26]
CuiZ L, CarterT E, BurtonJ W, Genetic base of 651 Chinese soybean cultivars released during 1923 to 1995. of 651 Chinese soybean cultivars released during 1923 to 1995. Crop Sci, 2000, 40: 1470-1481[本文引用:1][JCR: 1.513]
[27]
LiuK, MuseS V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics, 2005, 21: 2128-2129[本文引用:1][JCR: 5.323]
[28]
NeiM, TajimaF, TatenoY. Accuracy of estimated phylogenetic trees from molecular data. J Mol Evol, 1983, 19: 153-170[本文引用:1][JCR: 2.145]
[29]
盖钧镒, 赵团结, 崔章林, 邱家驯. 中国大豆育成品种中不同地理来源种质的遗传贡献. 中国农业科学, 1998, 31: 35-43GaiJ Y, ZhaoT J, CuiZ L, QiuJ X. Nuclear and cytoplasmic contributions of germplasm from distinct areas to the soybean cultivars released during 1923-1995 in China. Sci AgricSin, 1998, 31: 35-43 (in Chinese with English abstract)[本文引用:1][JCR: 1.44]
[30]
BarrettB A, KidwellK K, FoxP N. Comparison of AFLP and pedigree-gased genetic diversity assessment methods uing wheat cultivars from the Pacific Northwest. Crop Sci, 1998, 38: 1271-1278[本文引用:1][JCR: 1.513]


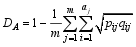

{kind=link}
{kind=link}