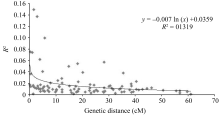
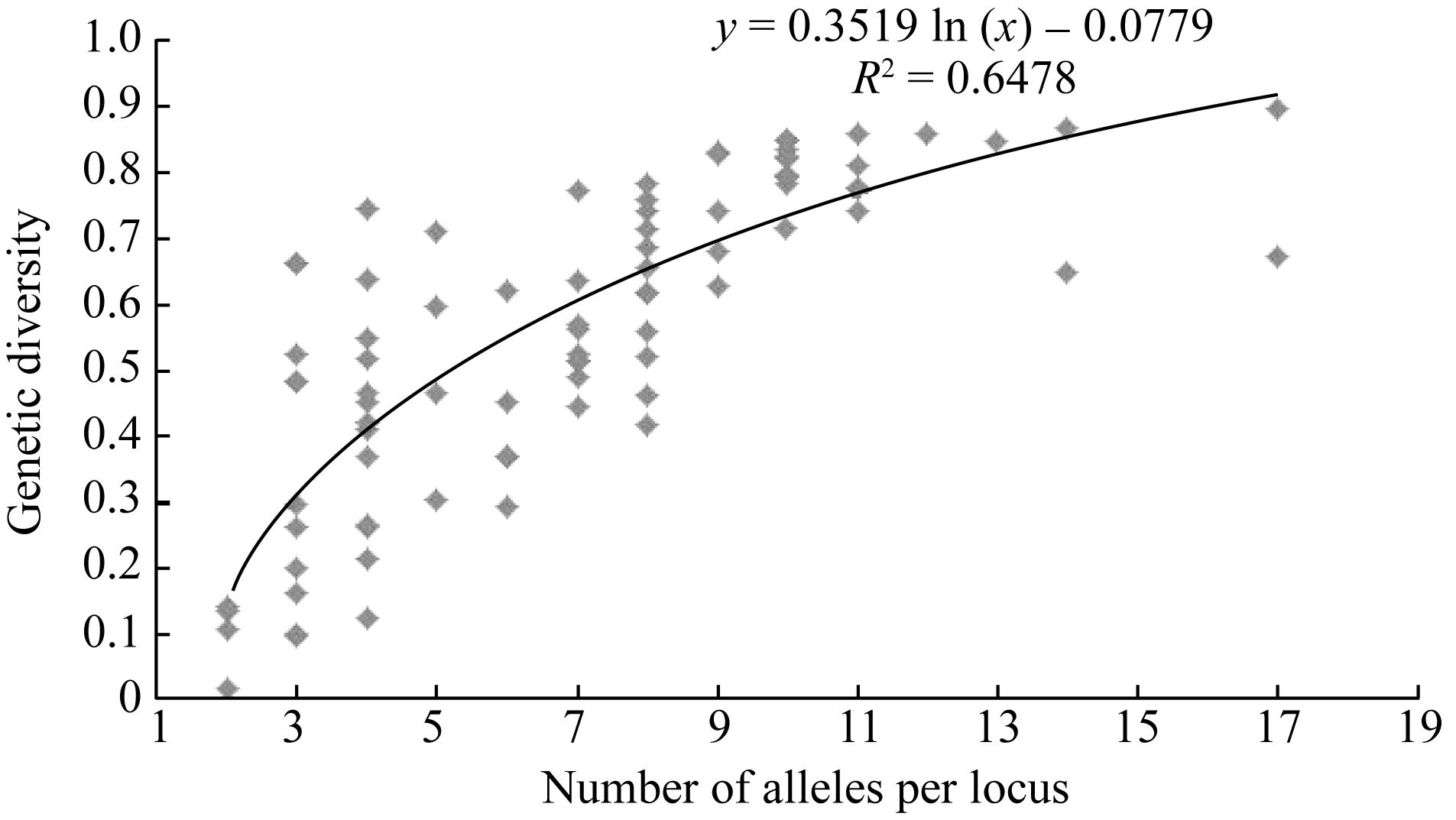
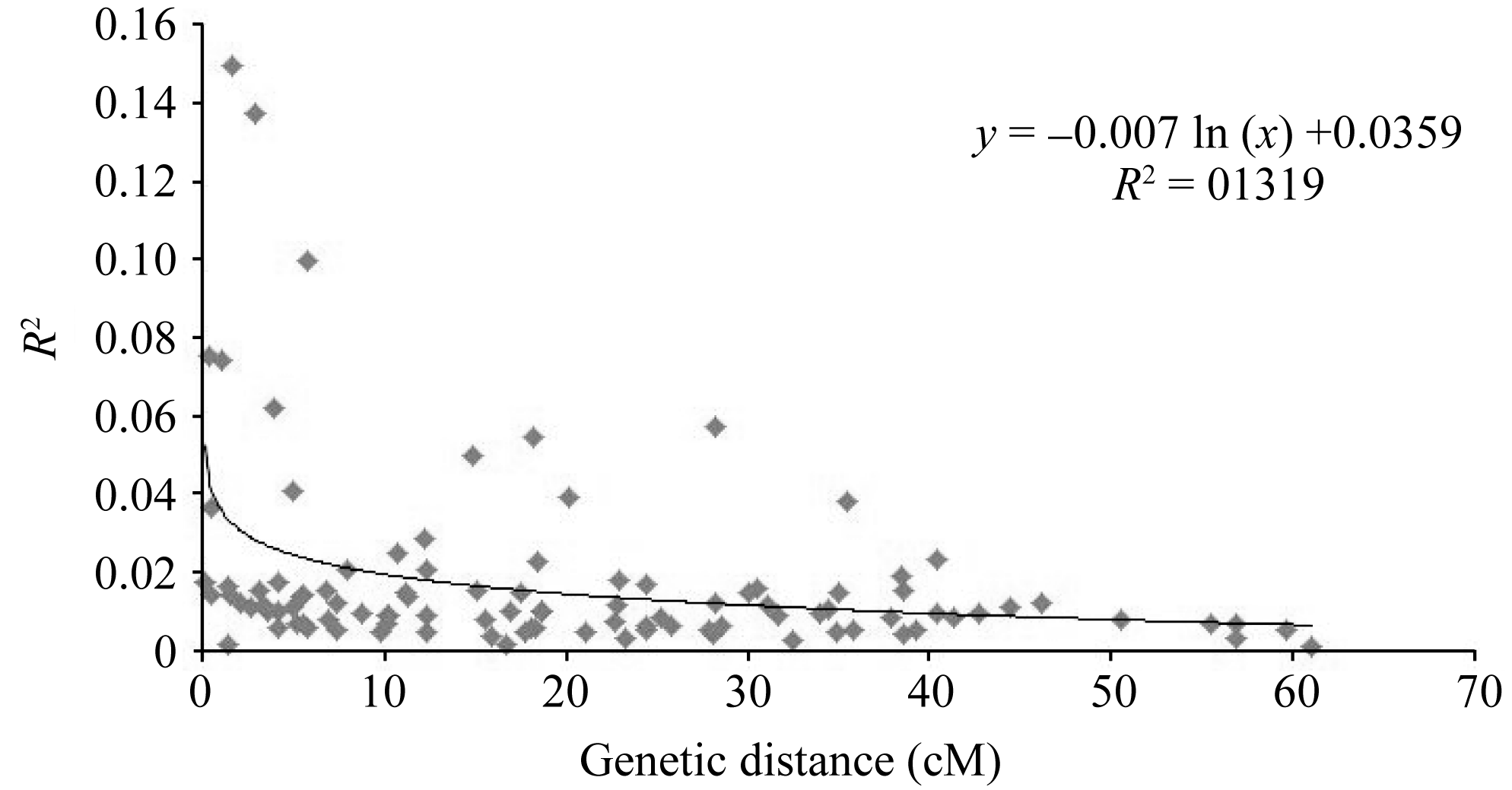
摘要利用57对小豆SSR标记和31对绿豆SSR标记, 用5份日本材料作对照, 对249份中国小豆种质进行遗传差异、群体结构和连锁不平衡(LD)分析。结果表明, 共检测到630个等位变异, SSR位点等位变异数在2~17之间, 遗传多样性指数范围为0.024~0.898, 平均为0.574。15个不同地理来源群体间表现出显著的遗传多样性差异, 其中中国云南最高, 河北和天津最低。聚类分析将254份材料划分为3个类群, 在一定程度上和地理生态环境相关。LD分析显示和其他作物相比, 小豆LD衰减距离较短, 最大衰减距离为5.8 cM (R2>0.1), 基因组LD平均衰减距离小于1 cM (R2>0.1,P<0.001)。
关键词:小豆; SSR标记; 遗传多样性; 群体结构; 连锁不平衡 Genetic Diversity, Population Structure and Linkage Disequilibrium in Adzuki Bean by Using SSR Markers BAI Peng, CHENG Xu-Zhen*, WANG Li-Xia, WANG Su-Hua, CHEN Hong-Lin Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Fund: AbstractA total of 249 Chinese adzuki bean accessions were analyzed with 57 adzuki bean SSR markers and 31 mungbean SSR markers to ascertain genetic diversity, population structure and linkage disequilibrium (LD) with five Japanese materials as the contrast. The results indicated that 630 alleles were detected with 2-17 alleles per locus and a mean genetic diversity index of 0.574, ranging from 0.024 to 0.898. There were significant differences among 15 populations of adzuki bean resources in genetic diversity from diversed geographic origins, with the highest in Yunnan and the lowest in Hebei and Tianjin. The 254 adzuki bean accessions could be divided into three subgroups based on STRUCTURE and NJ cluster. The population structure derived from them was positively correlated to some extent with the geographic eco-type. LD analysis revealed that there was a shorter LD decay distance in adzuki bean than that in other crops. The maximum LD decay distance, estimated by curvilinear regression, was 5.8 cM (R2>0.1), with a whole genome LD decay distance less than1 cM (R2>0.1,P<0.001). The results of this study should provide valuable information for future association mapping using this Chinese adzuki bean collection.
Keyword:Adzuki bean; SSR markers; Genetic diversity; Population structure; Linkage disequilibrium Show Figures Show Figures
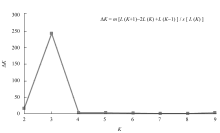
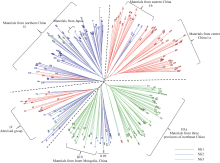
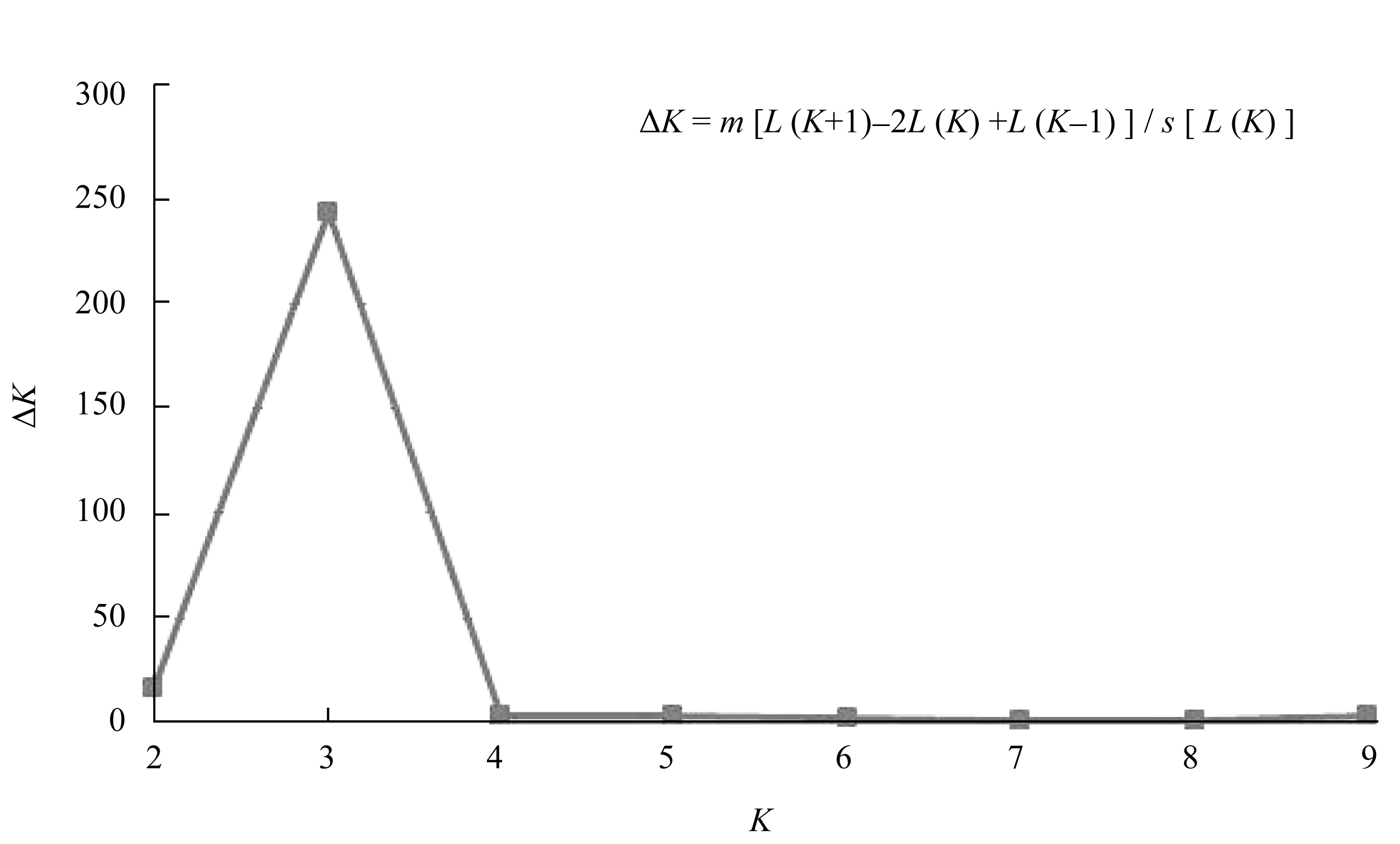
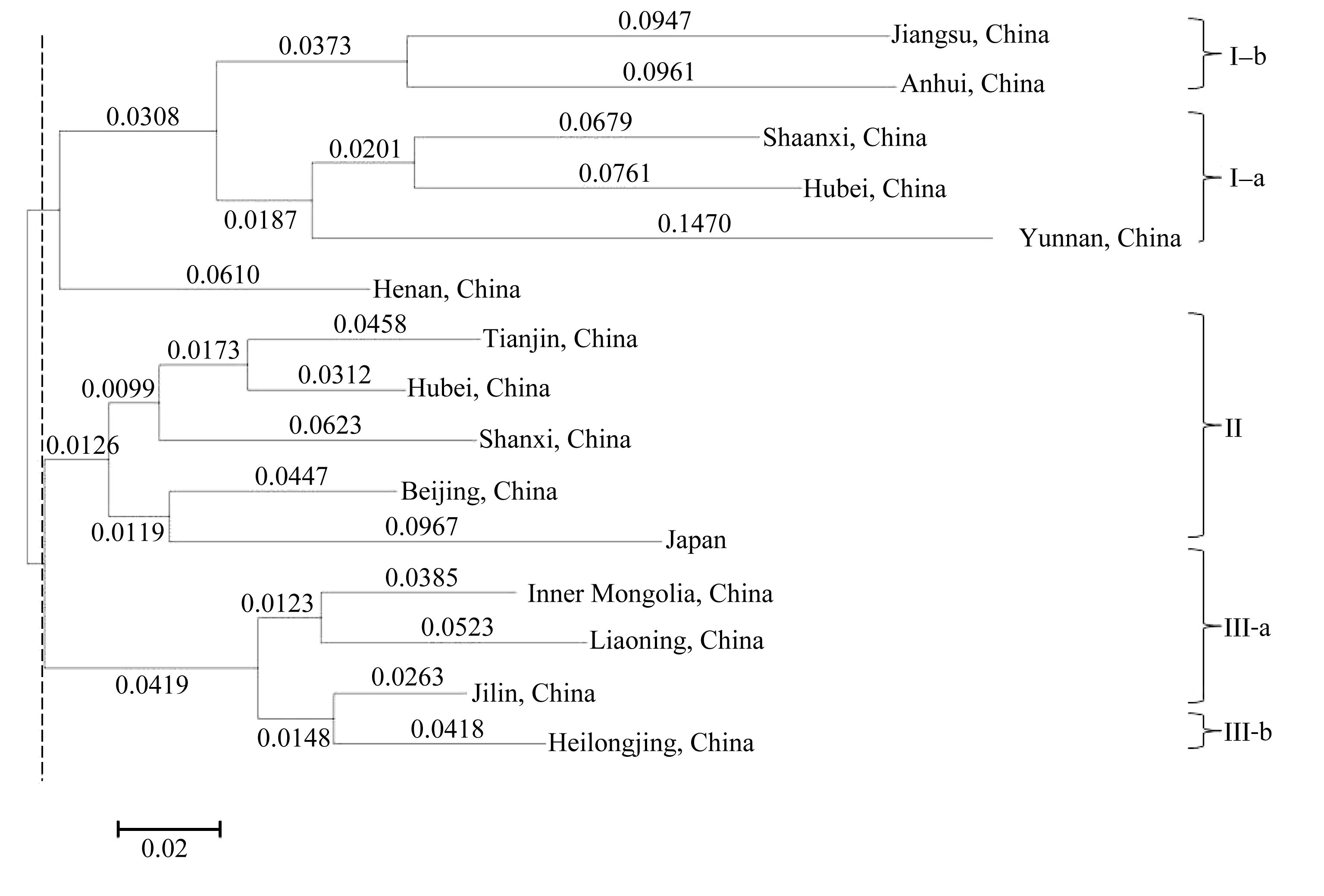
图3 基于NJ算法的254份小豆种质的系统树图SG1、SG2和SG3是由Structure在分群概率最大时分成的3个亚群。Fig. 3 Dendrogram of 254 adzuki bean accessions by NJ clusterSG1, SG2, and SG3 are three subgroups identified by Structure with the maximum membership probability.
4 结论本研究得到的我国小豆资源的遗传多样性水平较其他研究中的小豆种质要高。利用SSR标记进行Structure分析得到的群体结构要比使用地理生态型信息得到的更精确。小豆图谱上的共线性的或是非共性的SSR位点组合都有一定程度的LD。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
YoonM S, LeeJ, KimC Y, BaekH J. Genetic relationships among cultivated and wild Vigna angularis (Willd. ) Ohwi & Ohashi and relatives from Korea based on AFLP markers. Genet Resour Crop Evol, 2007, 54: 875-883[本文引用:1][JCR: 1.593]
[2]
金文林. 中国小豆生态气候资源分区初探. 北京农业科学, 1995, 13(6): 1-4JinW L. The Primary study on ecological regions of Chinese adzuki bean. Beijing Agric Sci, 1995, 13(6): 1-4 (in Chinese with English abstract)[本文引用:2]
[3]
ErlichH A, Gelfand D, SminskyJ J. Recent advances in polymerase chain reaction. Science, 1991, 252: 1643-1651[本文引用:1]
[4]
MullisK B. The Unusual origin of the polymerase chain reaction. Sci Am, 1990, 262: 56-61[本文引用:1][JCR: 1.478]
[5]
WangL X, GuanR X, LiuZ X, ChangR Z, QiuL J. Genetic diversity of Chinese cultivated soybean revealed by SSR markers. Crop Sci, 2006, 46: 1032-1038[本文引用:1][JCR: 1.513]
[6]
ChenX J, MinD H, TauqeerAhmad Yasir, HuY G. Genetic diversity, population structure and linkage disequilibrium in elite Chinese winter wheat investigated with SSR markers. PLoS One, 2012, 7: e44510[本文引用:1][JCR: 3.73]
[7]
Flint-GarciaS A, ThornsberryJ M, BucklerE S. Structure of linkage disequilibrium in plants. Annu Rev Plant Biol, 2003, 54: 357-374[本文引用:3][JCR: 25.962]
[8]
RafalskiA, MorganteM. Corn and humans: recombination and linkage disequilibrium in two genomes of similar size. Trends Genet, 2004, 20: 103-111[本文引用:1][JCR: 9.772]
[9]
ZhangX Y, TongY P, YouG X, HaoC Y, GeH M, WangL F, LiB, DongY C, LiZ S. Hitchhiking effect mapping: a new approach for discovering agronomic important genes. Sci Agric Sin, 2006, 39: 1526-1535[本文引用:1][CJCR: 1.889]
[10]
PritchardJ K, RosenbergN A. Use of unlinked genetic markers to detect population stratification in association studies. Am J Human Genet, 1999, 65: 220-228[本文引用:1][JCR: 11.202]
[11]
BucklerE S, ThornsberryJ M. Plant molecular diversity and applications to genomics. Plant Biol, 2002, 5: 107-111[本文引用:1][JCR: 2.32]
[12]
PritchardJ K, StephensM, DonnellyP. Inference of population structure using multilocus genotype data. Genetics, 2000, 155: 945-959[本文引用:1][JCR: 4.389]
[13]
RemingtonD L, ThornsberryJ M, MatsuokaY, WilsonL M, WhittS R, DoebleyJ, KresovichS, GoodmanM M, BucklerE S. Structure of linkage disequilibrium and phenotypic associations in the maize genome. Proc Natl Acad Sci USA, 2001, 98: 11479-11484[本文引用:2][JCR: 9.737]
[14]
KraakmanA T W, NiksR E, Van den BergP M M M, StamP, Van EeuwijkF A. Linkage disequilibrium mapping of yield and yield stability in modern spring barley cultivars. Genetics, 2004, 168: 435-446[本文引用:1][JCR: 4.389]
[15]
YuJ, PressoirG, BriggsW H, VrohBi I, YamasakiM, DoebleyJ F, McMullenM D, GautB S, NielsenD M, Holland J B, KresovichS, BucklerE S. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat Genet, 2006, 38: 203-208[本文引用:1][JCR: 35.209]
[16]
DoyleJ J, DoyleJ E. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull, 1987, 19: 11-15[本文引用:1]
[17]
HanO K, KagaA, IsemuraT, WangX W, TomookaN, VaughanD A. A genetic linkage map for adzuki bean [Vigna angularis (Willd. ) Ohwi & Ohashi]. Theor Appl Genet, 2005, 111: 1278-1287[本文引用:3][JCR: 3.658]
[18]
WangX X, KagaA, TomookaN, VaughanD A. The development of SSR markers by a new method in plants and their application to gene flow studies in adzuki bean [Vigna angularis (Willd. ) Ohwi & Ohashi]. Theor Appl Genet, 2004, 109: 352-360[本文引用:1][JCR: 3.658]
[19]
钟敏, 程须珍, 王丽侠, 王素华, 王小宝. 绿豆基因组SSR引物在豇豆属作物中的通用性. 作物学报, 2012, 38: 223-230ZhongM, ChengX Z, WangL X, WangS H, WangX B. Transferability of mungbean genomic-SSR markers in other Vigna species. Acta Agron Sin, 2012, 38: 223-230 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[20]
GoudetJ. FSTAT (Ver. 1. 2): a computer program to calculate F-statistics. J Hered, 1995, 86: 485-486[本文引用:1][JCR: 1.995]
[21]
RohlfF. NTSYSpc: Numerical Taxonomy System (Ver. 2. 2). Setauket, New York: Exeter Publishing, Ltd. 2006[本文引用:1]
[22]
YehF C, YangR C, BoyleT. Popgene Version 1. 31 Quick User Guide. University of Alberta, and Centre for International Forestry Research, Canada, 1999[本文引用:2]
[23]
LiuK, MuseS V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics, 2005, 21: 2128-2129[本文引用:1][JCR: 5.323]
[24]
EvannoG, RegnautS, GoudetJ. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol, 2005, 14: 2611-2620[本文引用:1][JCR: 6.275]
[25]
RambautA. FigTree. Ver. 1. 3. 1 [computer program]. [Issued on 2012-12-05][Visited on 2013-10-15]http://tree.bio.ed.ac.uk/[本文引用:1]
[26]
BradburyP J, ZhangZ, KroonD E, CasstevensT M, RamdossY, BucklerE S. TASSEL: software for association mapping of complex traits in diverse samples. Bioinform, 2007, 23: 2633-2635[本文引用:1]
[27]
濮绍京, 金文林, 赵波, 王占丽. 中国北方小豆地方品种资源研究. 北京农学院学报, 2003, 18: 174-177PuS J, JinW L, ZhaoB, WangZ L. Study on adzuki bean variety sources of North China. J Beijing Agric Coll, 2003, 18: 174-177 (in Chinese with English abstract)[本文引用:1]
[28]
金文林. 中国北方小豆地方品种资源的研究: I. 小豆农艺性状在南京生态条件下表现的相关性研究. 北京农学院学报, 1989, 4(3): 34-41JinW L. A research on local variety resources of adzuki bean of northern China: I. Study on correlation of quantitative traits of adzuki bean in Nanjing. J Beijing Agric Coll, 1989, (4): 34-41 (in Chinese with English abstract)[本文引用:1]
[29]
SharmaJ D, RathoreP K. Genetic divergence in adzuki bean. Crop Improv, 1994, 21: 49-53[本文引用:1]
[30]
ZongX X, KagaA, TomookaN, WangX W, HanO K, VaughanD. The genetic diversity of the Vigna angularis complex in Asia. Genome, 2003, 46: 647-658[本文引用:1][JCR: 1.668]
[31]
金文林, 文自翔, 濮绍京, 赵波. 应用RAPD分析小豆种质资源遗传多样性及遗传演化趋势. 中国农业科学, 2005, 38: 241-249JinW L, WenZ X, PuSJ, ZhaoB. Genetic diversity and evolution of adzuki bean germplasm resources based on RAPD markers. Sci Agric Sin, 2005, 38: 241-249 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[32]
叶剑, 赵波, 佟星, 濮绍京, 万平. 栽培小豆种质资源遗传多样性SSR标记分析. 北京农学院学报, 2008, 23: 8-13YeJ, ZhaoB, TongX, PuS J, WanP. Genetic diversity of cultivated adzuki bean germplasm resources based on SSR markers. J Beijing Univ Agric, 2008, 23: 8-13 (in Chinese with English abstract)[本文引用:1][CJCR: 0.6686]
[33]
王丽侠, 程须珍, 王素华. 利用SSR标记分析小豆种质资源的遗传多样性. 生物多样性, 2011, 19: 17-23WangL X, ChengX Z, WangS H. Genetic diversity in adzuki bean and its relatives based on SSR markers. Biodivers Sci, 2011, 19: 17-23 (in Chinese with English abstract)[本文引用:2]
[34]
KnowlerW C, WilliamsR C, PettittD J, SteinbergG A. Gin3;5, 13, 14 and type 2 diabetes mellitus: an association in American Indians with genetic admixture. Am J Hum Genet, 1988, 43: 520-526[本文引用:1][JCR: 11.202]
[35]
NordborgM, BorevitzJ O, BergelsonJ, BerryC C, ChoryJ, HagenbladJ, KreitmanM, MaloofJ N, NoyesT, OefnerP J, StahlE A, WeigelD. The extent of linkage disequilibrium in Arabidopsis thaliana. Nat Genet, 2002, 30: 190-193[本文引用:1][JCR: 35.209]
[36]
NordborgM, HuT T, IshinoY, JhaveriJ, ToomajianC, ZhengH G, CalabreseP, GoyalR, KimS, PadhukasahasramB, PlagnolV, RosenbergN A, ShahC, WallJ D, ZhaoK. The pattern of polymorphism in Arabidopsis thaliana. PloS Biol, 2005, 3: e196[本文引用:2][JCR: 12.69]
[37]
HytenD L, ChoiI Y, SongQ, ShoemakerR C, NelsonR L, CostaJ M, SpechtJ E, CreganP B. Highly variable patterns of linkage disequilibrium in multiple soybean populations. Genetics, 2007, 175: 1937-1944[本文引用:2][JCR: 4.389]
[38]
ParidaA, RainaS N, NarayanR K J. Quantitative DNA variation between and within chromosome complements of Vigna species (Fabaceae). Genetica, 1990, 82: 125-133[本文引用:1][JCR: 1.681]
[39]
徐宁, 程须珍, 王素华, 王丽侠, 赵丹. 以地理来源分组、利用表型数据构建中国小豆核心种质. 作物学报, 2008, 34: 1366-1373XuN, ChengX Z, WangS H, WangL X, ZhaoD. The establishment of Chinese adzuki bean core collection based on the geographical distribution and phenotypes. Acta Agron Sin, 2008, 34: 1366-1373 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[40]
王丽侠, 程须珍, 王素华, 徐宁, 梁辉, 赵丹. 利用SSR标记分析小豆种质资源的遗传多样性. 中国农业科学, 2009, 42: 2661-2666WangL X, ChengX Z, WangS H, XuN, LiangH, ZhaoD. Genetic diversity among adzuki bean germplasm revealed by SSR markers. Sci Agric Sin, 2009, 42: 2661-2666 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[41]
YeeE, KidwellK K, SillsG R, LumpkinT A. Diversity among selected Vigna angularis (Azuki) accessions on the basis of RAPD and AFLP markers. Crop Sci, 1999, 39: 268-275[本文引用:1][JCR: 1.513]
 , 王丽侠, 王素华, 陈红霖
, 王丽侠, 王素华, 陈红霖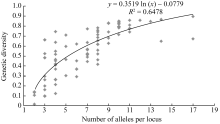


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}