关键词:甘蔗; S-腺苷甲硫氨酸合成酶; 克隆; 表达分析 Cloning and Expression of SugarcaneS-adenosylmethionine Synthetase GeneScSAM SONG Xiu-Peng1, ZHANG Bao-Qing2, HUANG Xing2, YANG Li-Tao1,2,*, LI Yang-Rui1,2,* 1Agricultural College / State Key Laboratory of Conservation and Utilization of Subtropical Agro-bioresources, Guangxi University, Nanning 530004, China
2Guangxi Academy of Agricultural Sciences / Sugarcane Research Center, Chinese Academy of Agricultural Sciences / Key Laboratory of Sugarcane Biotechnology and Genetic Improvement (Guangxi), Ministry of Agriculture / Guangxi Key Laboratory of Sugarcane Genetic Improvement, Nanning 530007, China
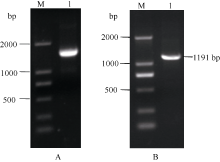
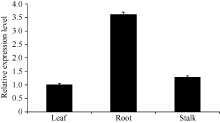
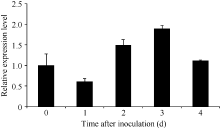
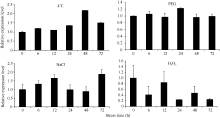
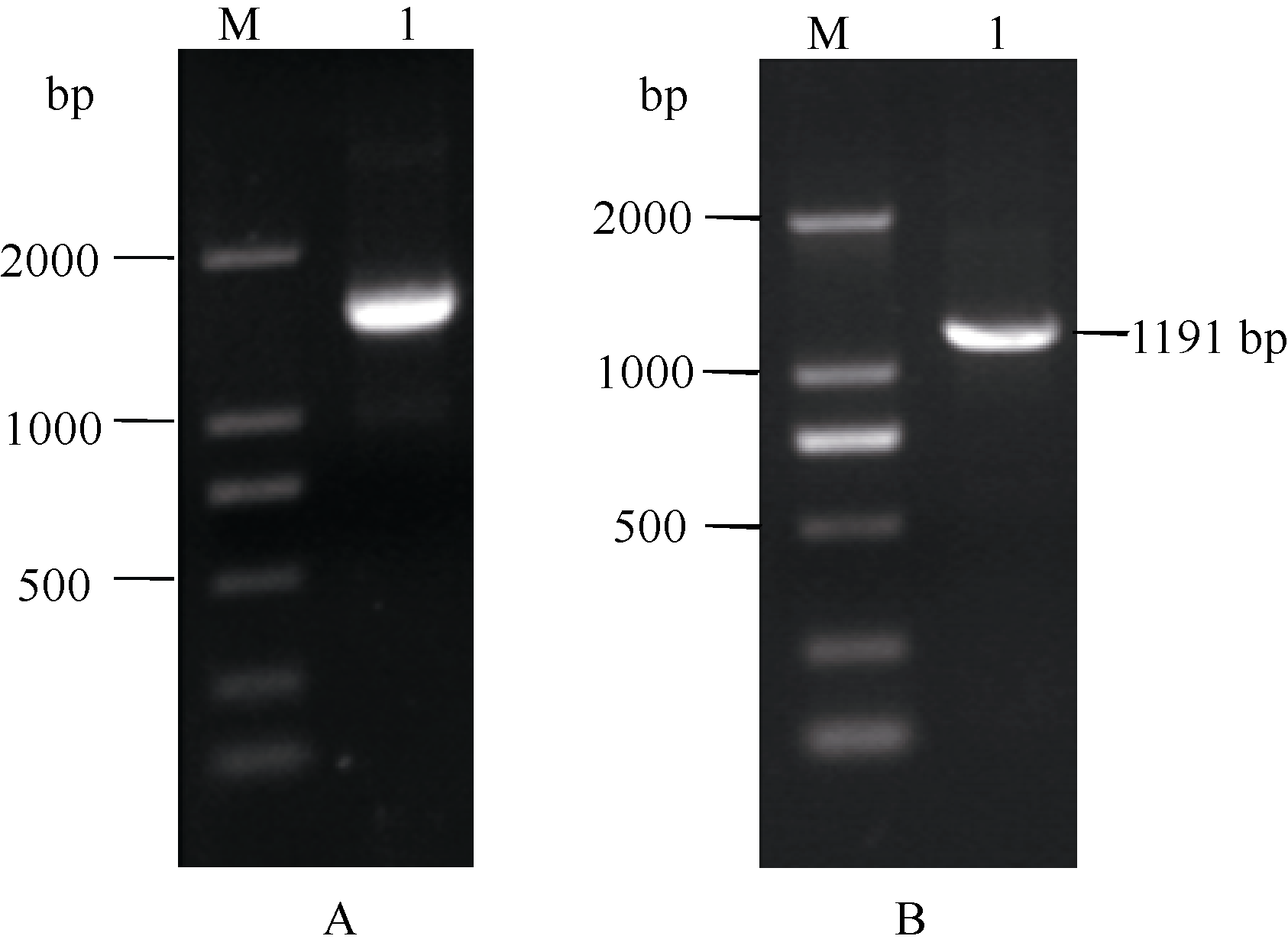
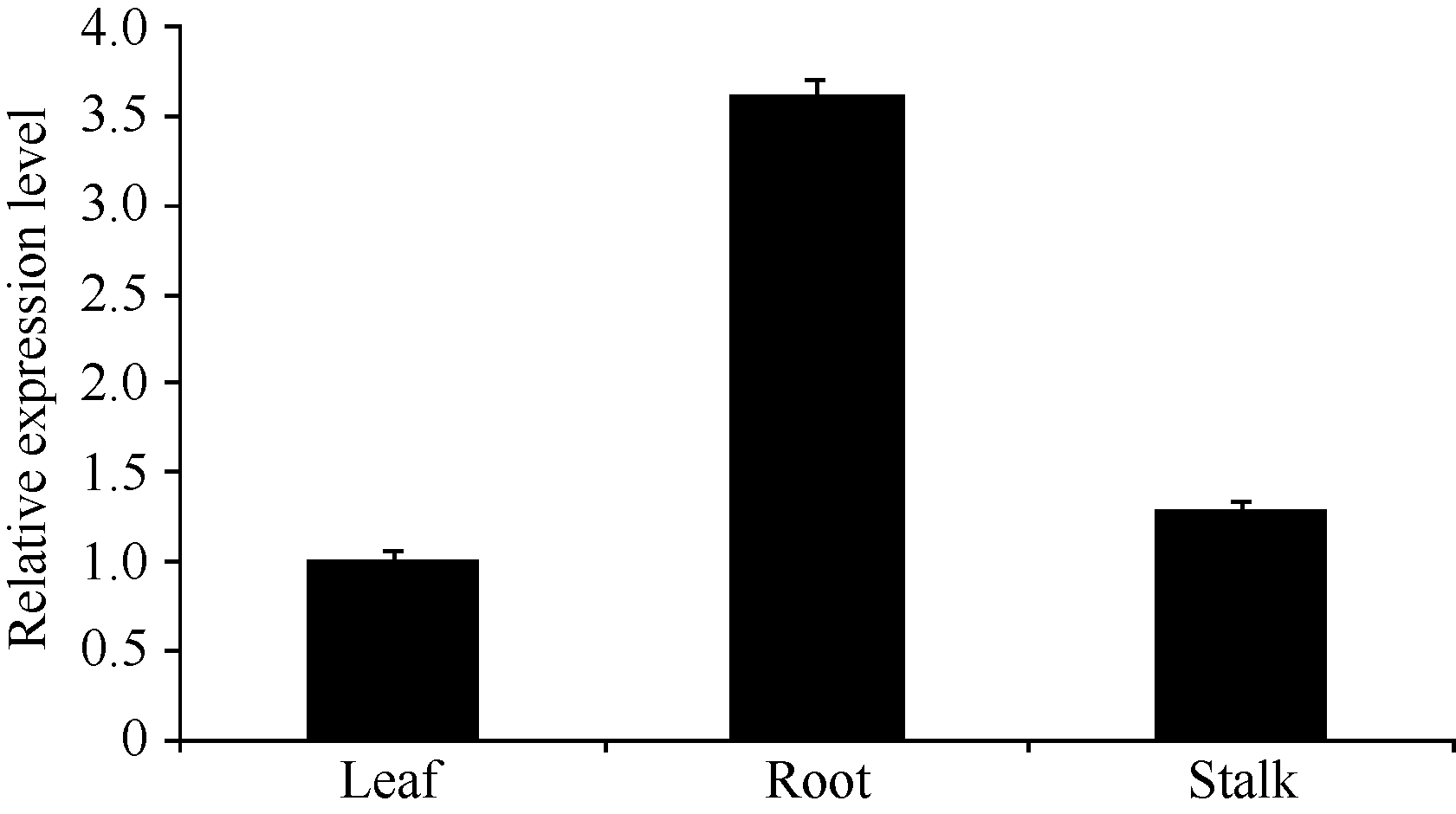
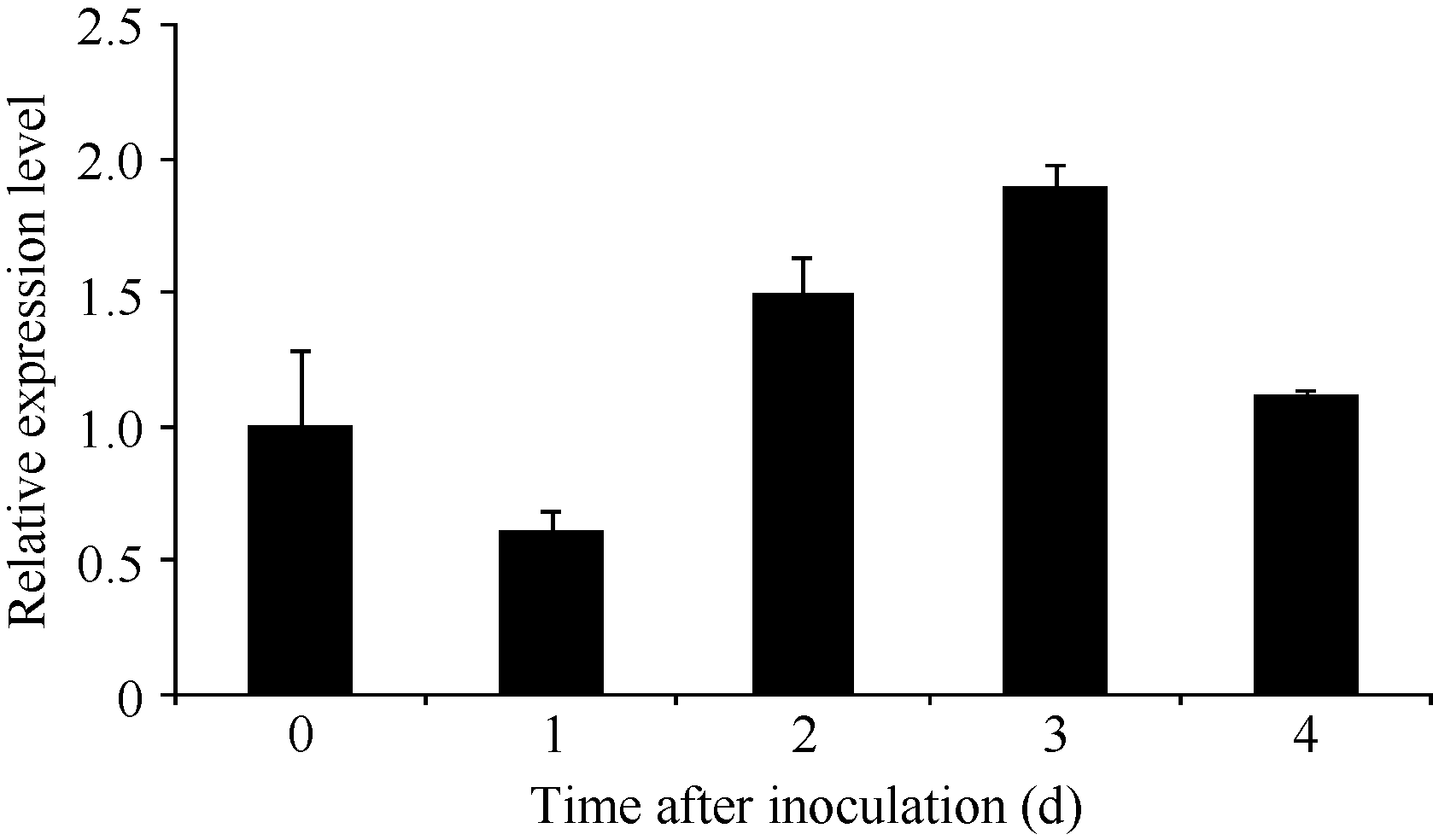
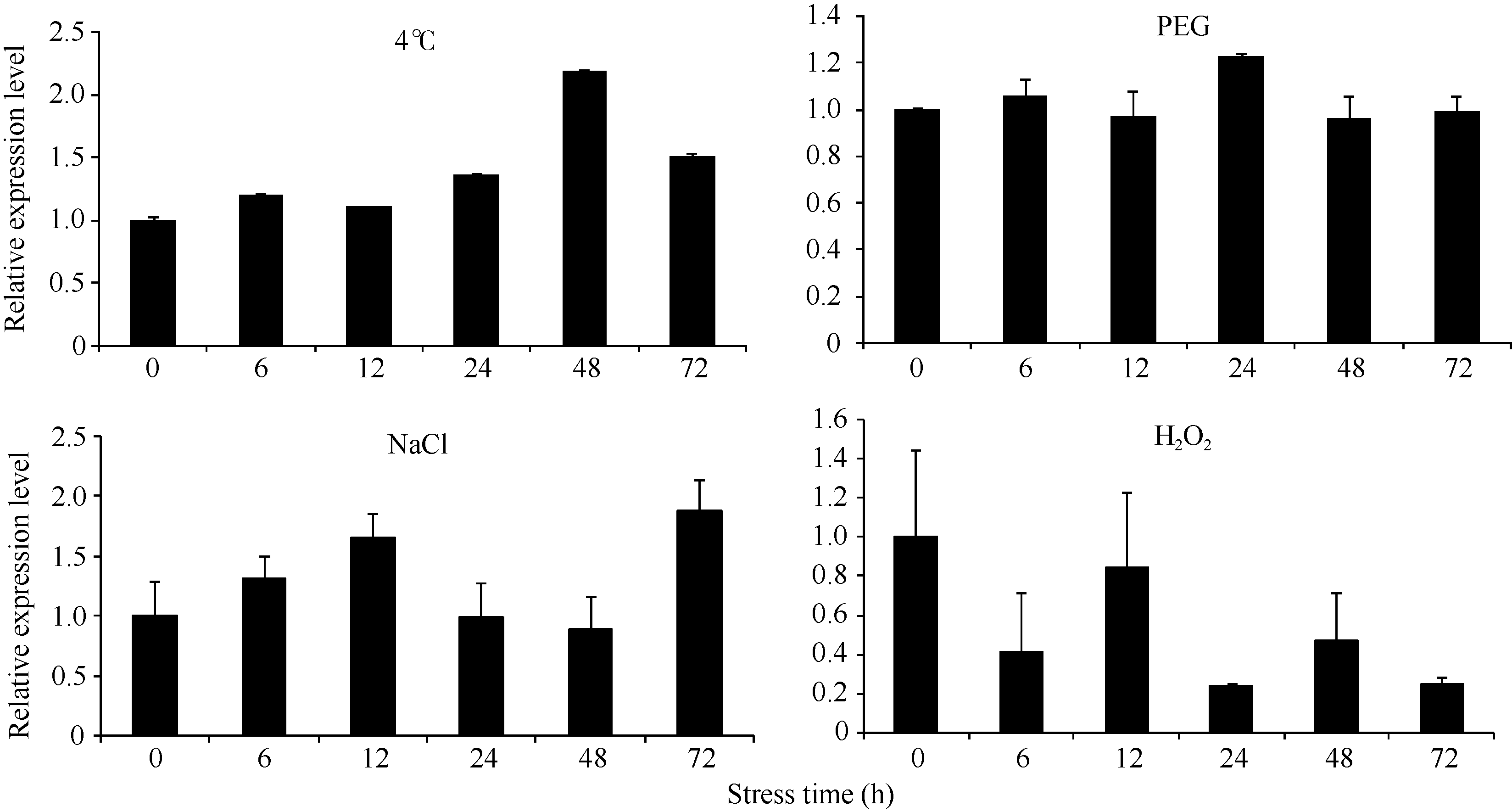
Fund: AbstractThe full-length sequence cDNA ofScSAM (GenBank accession number: KC172558) was cloned from sugarcane variety ROC 22 using RT-PCR combined with RACE techniques. This sequence consists of 1466 bp with an intact open reading frame of 1191 bp, encoding a polypeptide of 396 amino acids. Homology analysis showed that the deduced ScSAM protein was highly homologous to SAM proteins from different species. Phylogenetic analysis indicated thatScSAM was closely related to theSAM of sorghum. Real-time PCR results showed that theScSAM gene constitutively expressed in plant, with different expression levels in root, stalk and leaf. The transcript ofScSAMin root was the highest among the three organs, which was 3.6 times higher than that in leaf. Furthermore,ScSAM transcription was induced by biotic (smut infection) and abiotic (low temperature, PEG and NaCl) stresses, but the expression patterns were different. Under oxidative stress (H2O2), the expression ofScSAM was inhibited. We suggested thatScSAM might participate in smut-resistant activities in sugarcane, and also play a role in sugarcane resistances to chill, drought, salt and oxidation stresses.
Keyword:Sugarcane; S-adenosylmethionine synthetase (SAM); Clone; Expression analysis Show Figures Show Figures
图3 ScSAM编码区核苷酸序列及推导出的氨基酸序列起始密码子ATG和终止密码子TAA用下画线表示, S-腺苷甲硫氨酸合成酶特征序列为灰色。Fig. 3 Nucleotide and predicted amino acid sequences of ScSAMThe start codon (ATG) and stop codon (TAA) are underlined. The characteristic sequence of SAM is shaded by grey.
图5 ScSAM与其他植物SAM氨基酸序列的系统进化树各节点处数值表示BootStrap值(重复1000次)。Fig. 5 Phylogentic relationship of amino acid sequences between ScSAM and other SAM proteinsThe numbers next to the nodes give BootStrap values of 1000 replicates.
李杨瑞, 杨丽涛. 20世纪90年代以来我国甘蔗产业和科技的新发展. 西南农业学报, 2009, 22: 1469-1476LiY R. Yang L T. New developments of sugarcane industry and technology in China since 1990s. Southwest China J Agric Sci, 2009, 22: 1469-1476 (in Chinese with English abstract)[本文引用:1]
[2]
KumarT, Uzma, KhanM R, AbbasZ, AliG M. Genetic improvement of sugarcane for drought and salinity stress tolerance using Arabidopsis vacuolar pyrophosphatase (AVP1) gene. Mol Biotechnol, 2014, 56: 199-209[本文引用:1][JCR: 2.262]
[3]
MatoJ, AlvarezL, OrtizP, PajaresM A. S-adenosylmethionine synthesis: molecular mechanisms and clinical implications. Pharmacol Therapeutics, 1997, 73: 265-280[本文引用:1][JCR: 7.793]
BelbahriL, ChevalierL, BensaddekL, GilletF, FliniauxM A, BoerjanW, InzéD, ThomasD, ThomassetB. Different expression of an S-adenosylmethionine synthetase gene in transgenic tobacco callus modifies alkaloid biosynthesis. Biotechnol Bioeng, 2000, 69: 11-20[本文引用:2][JCR: 3.648]
[6]
GrundyF J, HenkinT M. Regulation of gene expression by effectors that bind to RNA. Curr Opin Microbiol, 2004, 7: 126-131[本文引用:1][JCR: 8.23]
[7]
WinklerW C, NahviA, SudarsanN, BarrickJ E, BreakerR R. An mRNA structure that controls gene expression by binding S-adenosylmethionine. Nat Struct Mol Biol, 2003, 10: 701-707[本文引用:2][JCR: 11.902]
[8]
KevbrinV V, ZenglerK, LysenkoA M, WiegelJ. Anoxybacillus kamchatkensis sp. nov. , a novel thermophilic facultative aerobic bacterium with a broad pH optimum from the Geyser valley, Kamchatka. Extremophiles, 2005, 9: 391-398[本文引用:1][JCR: 2.203]
EsparteroJ, Pintor-ToroJ A, PardoJ M. Differential accumulation of S-adenosylmethionine synthetase transcripts in response to salt stress. Plant Mol Biol, 1994, 25: 217-227[本文引用:1][JCR: 3.518]
[11]
MaX L, WangZ L, QiY C, ZhaoY X, ZhangH. Isolation of S-adenosylmethionine synthetase gene from Suaeda salsa and its differential expression under NaCl stress. Acta Bot Sin, 2003, 45: 1359-1365[本文引用:1][JCR: 0.599]
[12]
朱晶莹, 王寒玉, 张晏萌, 余爱丽. 玉米S-腺苷甲硫氨酸合成酶基因家族成员在盐胁迫条件下的差异表达. 核农学报, 2011, 25: 427-431ZhuJ Y, WangH Y, ZhangY M, YuA L. Differential expression of maize S-adenosylmethionin synthetase gene family members during salt stress. J Nucl Agric Sci, 2011, 25: 427-431 (in Chinese with English abstract)[本文引用:2][CJCR: 1.237]
[13]
LiX D, XiaB, WangR, XuS, JiangY M, YuF B, PengF. Molecular cloning and characterization of S-adenosylmethionine synthetase gene from Lycoris radiata. Mol Biol Rep, 2013, 40: 1255-1263[本文引用:1][JCR: 2.506]
[14]
樊金萍, 柏锡, 李勇, 纪巍, 王希, 才华, 朱延明. 野生大豆S-腺苷甲硫氨酸合成酶基因的克隆及功能分析. 作物学报, 2008, 34: 1581-1587FanJ P, BoX, LiY, JiW, WangX, CaiH, ZhuY M. Cloning and function analysis of gene SAMS from Glycine soja. Acta Agron Sin, 2008, 34: 1581-1587 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[15]
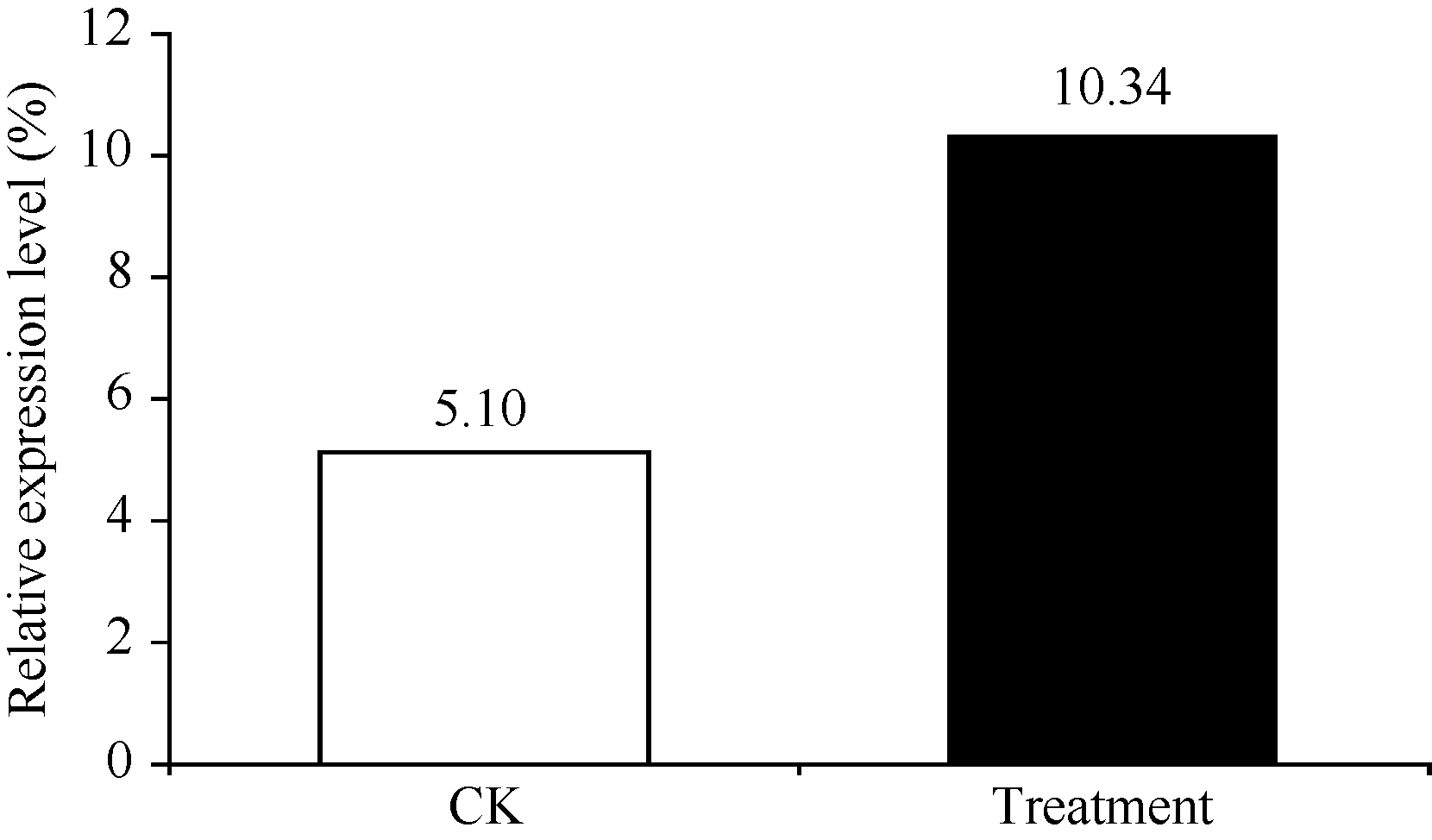
SantiagoR, deArmas R, FontaniellaB, VicenteC, LegazM E. Changes in soluble and cell wall-bound hydroxycinnamic and hydroxybenzoic acids in sugarcane cultivars inoculated with Sporisorium scitamineum sporidia. Eur J Plant Pathol, 2009, 124: 439-450[本文引用:1][JCR: 1.61]
[16]
宋修鹏, 黄杏, 莫凤连, 杨丽涛, 李杨瑞, 陈保善. 甘蔗苯丙氨酸解氨酶基因(PAL)的克隆和表达分析. 中国农业科学, 2013, 46: 2856-2868SongX P, HangX, MoF L, YangL T, LiY R, ChenB S. Cloning and expression analysis of sugarcane phenylalanin ammonia- lyase (PAL) gene. Sci Agric Sin, 2013, 46: 2856-2868 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[17]
CantoniG. The nature of the active methyl donor formed enzymatically from L-methionine and adenosinetriphosphate. J Am Chem Soc, 1952, 74: 2942-2943[本文引用:1][JCR: 10.677]
[18]
柴兴苹, 张玉秀, 谭金娟, 冯珊珊, 柴团耀. Zn胁迫下小麦S-腺苷甲硫氨酸代谢途径关键基因表达模式分析. 植物生理学报, 2013, 49: 375-384ChaiX P, ZhangY X, TanJ J, FengS S, ChaiT Y. Analysis of expression patterns of genes participated in S-adenosylme-thionine (SAM) metabolic pathway in wheat under Zn stress. Plant Physiol J, 2013, 49: 375-384 (in Chinese with English abstract)[本文引用:1][JCR: 2.699]
[19]
KasukabeY, HeL, NadaK, MisawaS, IharaI, TachibanaS. Overexpression of spermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stresS-regulated genes in transgenic Arabidopsis thaliana. Plant Cell Physiol, 2004, 45: 712-722[本文引用:1][JCR: 4.134]
[20]
陈坤明, 张承烈. 干旱期间春小麦叶片多胺含量与作物抗旱性的关系. 植物生理学报, 2000, 26: 381-386ChenK M, ZhangC L. Polyamine contents in the spring wheat leaves and their relations to drought-resistance. Plant Physiol J, 2000, 26: 381-386 (in Chinese with English abstract)[本文引用:1][JCR: 2.699]
[21]
鞠妍, 侯和胜. SAM合成酶基因的研究进展与应用现状. 天津农业科学, 2012, 18: 27-29JuY, HouH S. Research and application status of SAM synthase gene. Tianjin Agric Sci, 2012, 18: 27-29 (in Chinese with English abstract)[本文引用:1][CJCR: 0.5293]
[22]
周向红, 王萍. 向日葵S-腺苷甲硫氨酸合成酶基因克隆与分析. 作物杂志, 2011, (6): 10-13ZhouX H, WangP. Molecular cloning and bioinformatic analysis of S-adenosylmethionine synthetase gene from Helianthus annuus. Crops, 2011, (6): 10-13 (in Chinese with English abstract)[本文引用:1][CJCR: 0.6276]
[23]
孔令芳, 张吉宇, 刘志鹏, 王彦荣. 无芒隐子草SAMS1基因的克隆及干旱胁迫下的表达分析. 草业学报, 2013, 22: 268-275KongL F, ZhangJ Y, LiuZ P, WangY R. Cloning of a S-adenosyl methionine synthetase gene from Cleistogenes songorica and its expression under droght stress. Acta Pratac Sin, 2013, 22: 268-275 (in Chinese with English abstract)[本文引用:1]
[24]
JenningsJ C, Apel-BirkholdP C, MockN M, BakerC J, AndersonJ D, BaileyB A. Induction of defense responses in tobacco by the protein Nep1 from Fusarium oxysporum. Plant Sci, 2001, 161: 891-899[本文引用:1][JCR: 2.922]
[25]
KodakiT, TsujiS, OtaniN, YamamotoD, RaoK S, WatanabeS, TsukatsuneM, MakinoK. Differential transcriptional regulation of two distinct S-adenosylmethionine synthetase genes (SAM1 and SAM2) of Saccharomyces cerevisiae. Nucl Acids Res, 2003, 3(suppl): 303-304[本文引用:1]
[26]
MathurM, SharmaN, SacharR. Differential regulation of S-adenosylmethionine synthetase isozymes by gibberellic acid in dwarf pea epicotyls. Biochim Biophys Acta (BBA)-Protein Struct Mol Enzymol, 1993, 1162: 283-290[本文引用:1]
[27]
HazarikaP, RajamM V. Biotic and abiotic stress tolerance in transgenic tomatoes by constitutive expression of S-adenosylmethionine decarboxylase gene. Physiol Mol Biol Plants, 2011, 17: 115-128[本文引用:1][JCR: 25.962]
[28]
YueC W, XiaoJ, LingX, ZengN. Effect of low temperature stress on sweet potato S-adenosyl methionine synthetase gene expression. Agric Sci Technol, 2008, 9: 11-14[本文引用:1][CJCR: 2.0208]
[29]
陈锐, 陈亮, 王士强, 胡银岗. 水分胁迫下小麦S-腺苷甲硫氨酸合成酶基因的半定量表达模式分析. 麦类作物学报, 2009, 29: 954-958ChenR, ChenL, WangS Q, HuY G. Expression analysis of wheat S-adenosylmethionine synthetase gene during water stress and re-watering by semi-quantitative RT-PCR. J Triticeae Crops, 2009, 29: 954-958 (in Chinese with English abstract)[本文引用:2]
[30]
林凡云, 王士强, 胡银岗, 何蓓如. 糜子SAMS基因的克隆及其在干旱复水中的表达模式分析. 作物学报, 2008, 34: 777-782LinF Y, WangS Q, HuY G, HeB R. Cloning of a S-adenosylmethionine synthetase gene from broomcorn millet (Panicum miliaceum L. ) and its expression during drought and re-watering. Acta Agron Sin, 2008, 34: 777-782 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[31]
周凯, 宋丽艳, 叶武威, 王俊娟, 王德龙, 樊保香. 陆地棉耐盐相关基因GhSAMS的克隆及表达. 作物学报, 2011, 37: 1012-1019ZhouK, SongL Y, YeW W, WangJ J, WangD L, FanB X. Cloning and expression of GhSAMS gene related to salt-tolerance in Gossypium hirsutum L. Acta Agron Sin, 2011, 37: 1012-1019 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[32]
李燕, 孙伟, 赵彦修, 张慧. 盐地碱蓬甲硫氨酸合成酶基因(SsMS)的克隆与表达分析. 山东师范大学学报(自然科学版), 2003, 18: 76-78LiY, SunW, ZhaoY X, ZhangH. Isolation and characteizing of a methionine synthetase gene in Suaeda salsa under stress. J Shand ong Norm Univ (Nat Sci), 2003, 18: 76-78 (in Chinese with English abstract)[本文引用:1]
[33]
Sánchez-AguayoI, Rodríguez-GalánJ M, GarcíaR, TorreblancaJ, PardoJ M. Salt stress enhances xylem development and expression of S-adenosyl-L-methionine synthase in lignifying tissues of tomato plants. Planta, 2004, 220: 278-285[本文引用:1][JCR: 3.347]
 , 张保青
, 张保青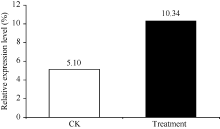







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}