关键词:棉花; TRV-VIGS; GhCLA1; GhMAPKKK; 黄萎病 Establishment and Application of TRV-Mediated Virus-Induced Gene Silencing in Cotton WANG Xin-Yu1,*, LÜ Kun1, CAI Cai-Ping2, XU Jun2, GUO Wang-Zhen2 1 College of Life Sciences, Nanjing Agricultural University, Nanjing 210095, China
2 State Key Laboratory of Crop Genetics & Germplasm Enhancement / College of Agriculture, Nanjing Agricultural University, Nanjing 210095, China
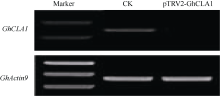
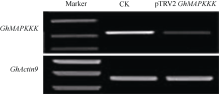
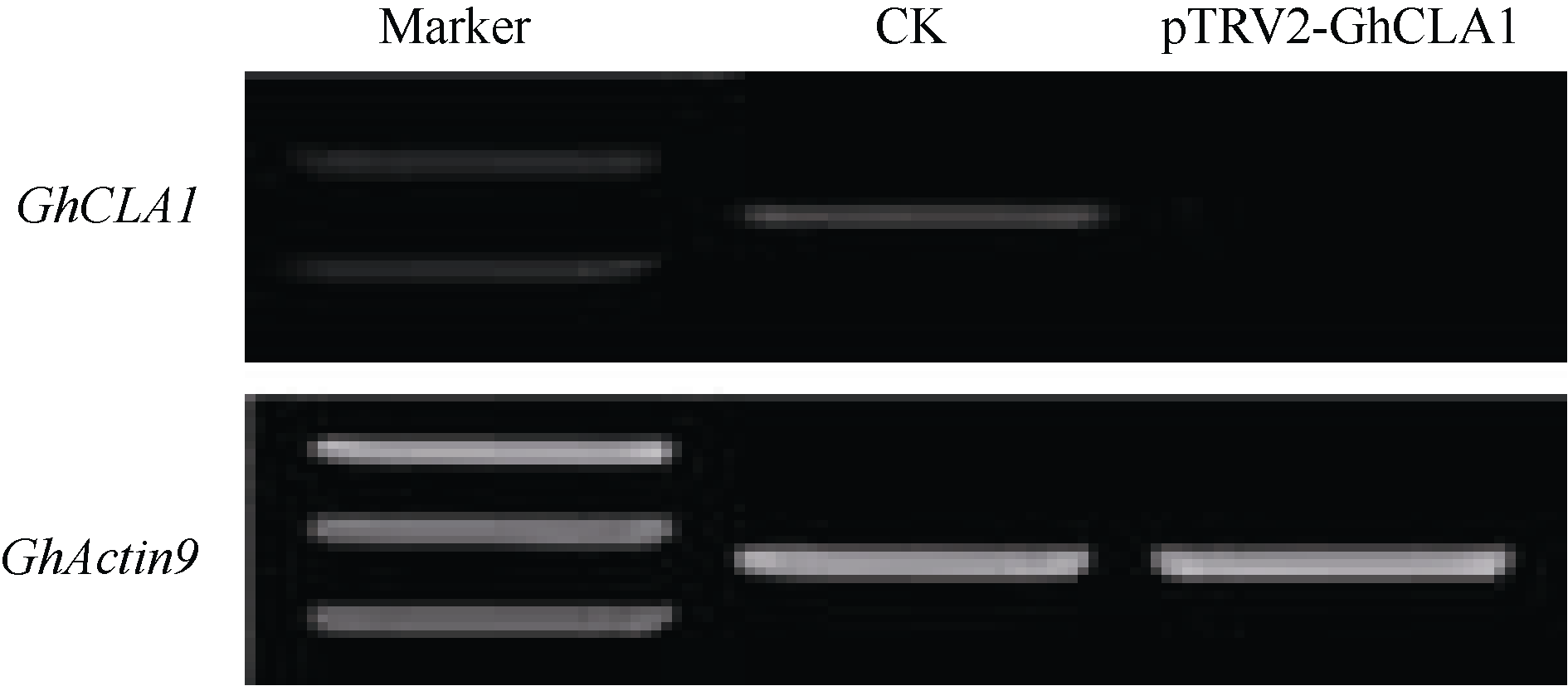
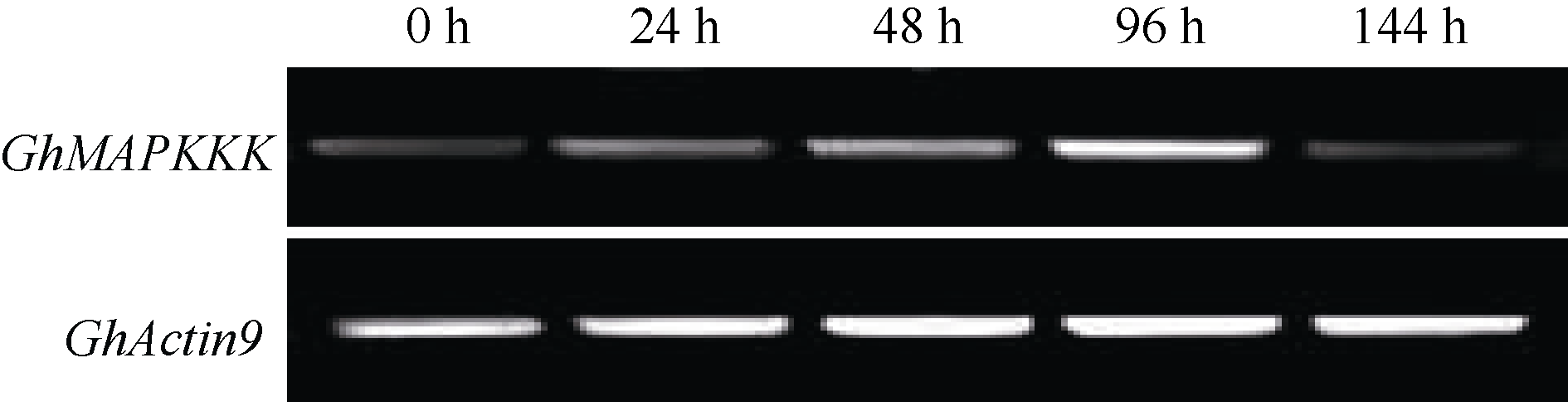
Fund: AbstractUsing upland cottonGhCLA1 as marker gene, tobacco rattle virus induced genes silencing (TRV-VIGS) was established in cotton. A gene fragment of TRV subgenomic RNA2 was amplified by reverse transcription-polymerase chain reaction (RT-PCR) in root, stem and leaf of cotton, demonstrating the virus can spread into various organs. The TRV induced silencing ofCLA1 gene was further tested in 34 different cotton varieties (lines) originated from several ecological regions of China. The results showed thatCLA1 could be silenced in all tested varieties (lines), though the levels of silencing showed a little difference, implicating wide application perspective of TRV-VIGS system in cotton.GhMAPKKK, a mitogen-activated protein kinase (MAPK) kinase kinase gene of cotton, was up-regulated in cotton at 96 hours post inoculation byVerticillium dahliae. SilencingGhMAPKKK inG. barbadense cv. Hai 7124, a cotton variety with high resistance toV. dahlia exhibited reduced resistance toV. dahliaeinfection, suggesting thatGhMAPKKK participated in cotton resistance signaling pathway toV. dahliae. With the wide adaption without genotype selection, sensitivity and high throughput, the TVR-VIGS system will significantly promote functional gene analysis in cotton.
图1 沉默棉花 GhCLA1的白化表型A: 整株表型, 左为接种pTRV1和pTRV2空载体对照, 右为接种pTRV1和pTRV2-GhCLA1植株的真叶表型。B: 叶片沉默效果。从左到右: 第一片真叶到第四片真叶。Fig. 1 Albino phenotype of GhCLA1silenced cotton plantsA: phenotype of cotton plant, the left is a control plant inoculated with pTRV1/pTRV2 constructs, the right is a cotton plant inoculated with pTRV1/pTRV2-GhCLA1 constructs; B: phenotype of GhCLA1 silenced cotton leaves; from left to right: the first to the fourth true leaves.
图4 34份不同来源棉花品种/品系TRV-VIGS诱导 GhCLA1基因沉默表型编号对应的材料见表1。每一材料上方为注射空载体对照, 下方为沉默 GhCLA1基因的叶片。Fig. 4 Photobleaching phenotype of GhCLA1silenced cotton plants in 34 cotton varieties (lines)Code numbers of cotton varieties correspond with these in Table 1. In each column, the upper is the leaves of vector control plants and the lower is those of GhCLA1 silenced plants.
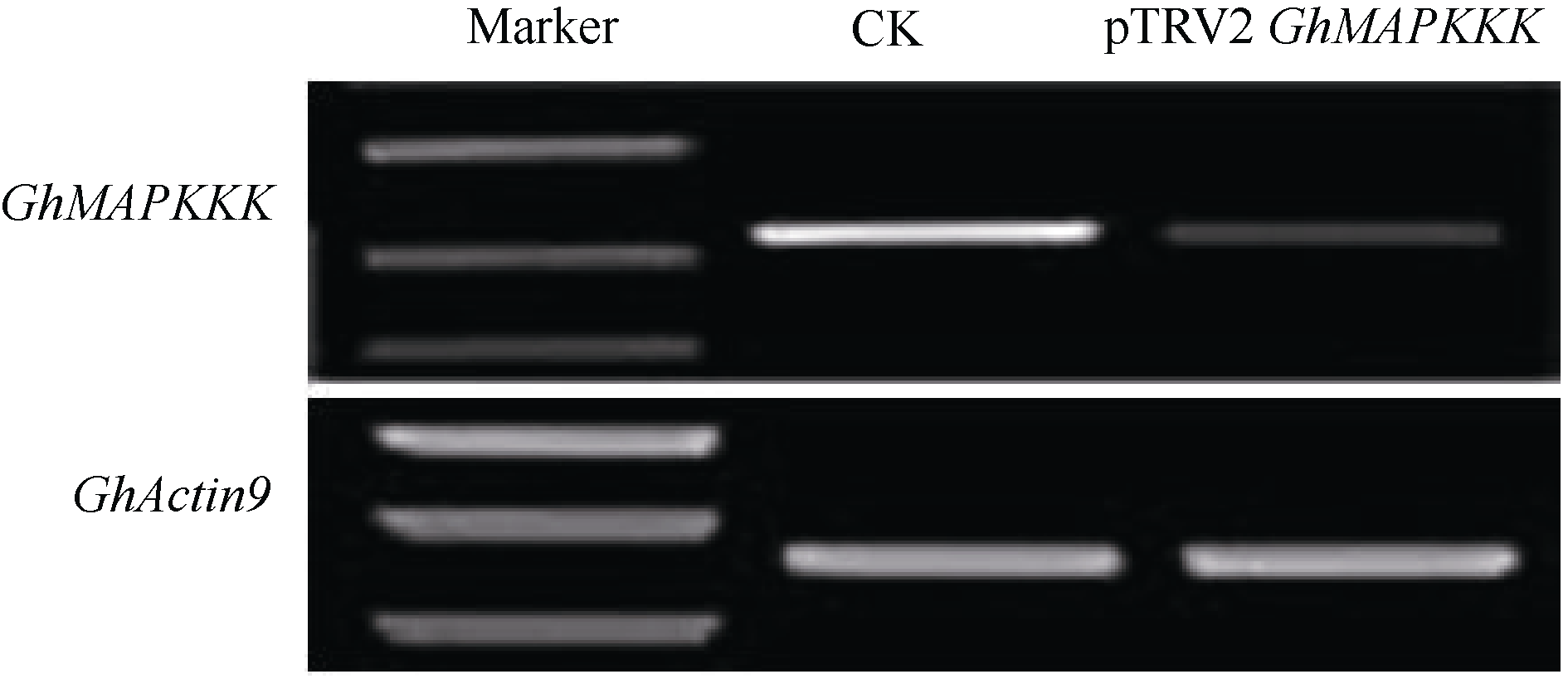
图7 黄萎病菌侵染 GhMAPKKK基因沉默后的海7124表型上排和下排4株分别为 GhMAPKKK 沉默及注射空载体对照后接种黄萎病菌的表型。Fig. 7 Wilt symptom of GhMAPKKK silenced Hai 7124 plants inoculated with Verticillium dahliae strain Vd8Four plants on the upper and lower respectively: disease symptoms in GhMAPKKK silenced and vector control plants in response to Verticillium dahliainfection.
LuR, Martin-Hernand ezA M, PeartJ R. Virus-induced gene silencing in plant. Methods, 2003, 30: 296-303[本文引用:1][JCR: 3.641]
[2]
PurkayasthaA, DasguptaI. Virus-induced gene silencing: a versatile tool for discovery of gene functions in plants. Plant Physiol Biochem, 2009, 47: 967-976[本文引用:1]
[3]
RyuC M, Anand A, KangL, MysoreK S. Agrodrench: a novel and effective agroinoculation method for virus-induced gene silencing in roots and diverse Solanaceous species. Plant J, 2004, 40: 322-331[本文引用:1][JCR: 6.582]
[4]
Senthil-KumarM, HemaR, Anand A, KangL, UdayakumarM, MysoreK S. A systematic study to determine the extent of gene silencing in Nicotiana benthamiana and other Solanaceae species when heterologous gene sequences are used for virus- induced gene silencing. New Phytol, 2007, 176: 782-791[本文引用:1][JCR: 6.736]
[5]
CzosnekH, EybishtzA, SadeD, GorovitsR, SobolI, BejaranoE, Rosas-DíazT, Lozano-DuránR. Discovering host genes involved in the infection by the tomato yellow leaf curl virus complex and in the establishment of resistance to the virus using tobacco rattle virus-based post transcriptional gene silencing. Viruses, 2013, 5: 998-1022[本文引用:1]
[6]
Burch-SmithT M, SchiffM, LiuY, Dinesh-KumarS P. Efficient virus-induced gene silencing in Arabidopsis. Plant Physiol, 2006, 142: 21-27[本文引用:1][JCR: 6.555]
[7]
YeJ, QuJ, BuiH T, ChuaN H. Rapid analysis of Jatropha curcas gene functions by virus-induced gene silencing. Plant Biotechnol J, 2009, 7: 964-976[本文引用:1][JCR: 6.279]
[8]
SpitzerB, ZviM M, OvadisM, MarhevkaE, BarkaiO, EdelbaumO, MartonI, MasciT, AlonM, MorinS, RogachevI, AharoniA, VainsteinA. Reverse genetics of floral scent: application of tobacco rattle virus-based gene silencing in Petunia. Plant Physiol, 2007, 145: 1241-1250[本文引用:1][JCR: 6.555]
[9]
HilemanL C, DreaS, MartinoG, LittA, IrishV F. Virus-induced gene silencing is an effective tool for assaying gene function in the basal eudicot species Papaver somniferum (opium poppy). Plant J, 2005, 44: 334-341[本文引用:1][JCR: 6.582]
[10]
WegeS, ScholzA, GleissbergS, BeckerA. Highly efficient virus-induced gene silencing (VIGS) in California poppy (Eschscholzia californica): an evaluation of VIGS as a strategy to obtain functional data from non-model plants. Ann Bot, 2007, 100: 641-649[本文引用:1][JCR: 0.657]
[11]
TuttleJ R, IdrisA M, BrownJ K, HaiglerC H, RobertsonD. Geminivirus-mediated gene silencing from cotton leaf crumple virus is enhanced by low temperature in cotton. Plant Physiol, 2008, 148: 41-50[本文引用:1][JCR: 6.555]
[12]
Mand elM A, FeldmannK A, Herrera-EstrellaL, Rocha-SosaM, LeónP. CLA1, a novel gene required for chloroplast development, is highly conserved in evolution. Plant J, 1996, 9: 649-658[本文引用:1][JCR: 6.582]
[13]
GaoX, Britt R C Jr, ShanL, HeP. Agrobacterium-mediated virus-induced gene silencing assay in cotton. J Vis Exp, 2011, 54: pii: 2938. DOI: DOI:10.3791/2938[本文引用:1]
[14]
GaoX, WheelerT, LiZ, KenerleyC M, HeP, ShanL. Silencing GhNDR1 and GhMKK2 compromises cotton resistance to Verticillium wilt. Plant J, 2011, 66: 293-305[本文引用:2][JCR: 6.582]
[15]
QuJ, YeJ, GengY F, SunY W, GaoS Q, ZhangB P, ChenW, ChuaN H. Dissecting functions of KATANIN and WRINKLED1 in cotton fiber development by virus-induced gene silencing. Plant Physiol, 2012, 160: 738-748[本文引用:3][JCR: 6.555]
[16]
PatersonA H, WendelJ F, GundlachH, GuoH, JenkinsJ, JinD, LlewellynD, ShowmakerK C, ShuS, UdallJ, YooM J, ByersR, ChenW, Doron-FaigenboimA, DukeM V, GongL, GrimwoodJ, GroverC, GruppK, HuG, LeeT H, LiJ, LinL, LiuT, MarlerB S, PageJ T, RobertsA W, RomanelE, Sand ersW S, SzadkowskiE, TanX, TangH, XuC, WangJ, WangZ, ZhangD, ZhangL, AshrafiH, BedonF, BowersJE, BrubakerC L, CheeP W, DasS, GingleA R, HaiglerC H, HarkerD, HoffmannL V, HovavR, JonesD C, LemkeC, MansoorS, urRahman M, RainvilleL N, RambaniA, ReddyU K, RongJ K, SarangaY, SchefflerB E, SchefflerJ A, StellyD M, TriplettB A, VanDeynze A, VaslinM F, WaghmareV N, WalfordS A, WrightR J, ZakiE A, ZhangT, DennisE S, MayerK F, PetersonD G, RokhsarD S, WangX, SchmutzJ. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature, 2012, 492: 423-427[本文引用:3][JCR: 38.597]
[17]
VelásquezA C, ChakravarthyS, MartinG B. Virus-induced gene silencing (VIGS) in Nicotiana benthamiana and tomato. J Vis Exp, 2009, 28: PII: 1292. DOI: DOI:10.3791/1292[本文引用:1]
[18]
蒋建雄, 张天真. 利用CTAB/酸酚法提取棉花组织总RNA. 棉花学报, 2003, 15: 166-167JiangJ X, ZhangT Z. Extraction of total RNA in cotton tissues with CTAB-acidic phenolic method. Cotton Sci, 2003, 15: 166-167[本文引用:1][CJCR: 1.462]
[19]
徐理, 朱龙付, 张献龙. 棉花抗黄萎病机制研究进展. 作物学报, 2012, 38: 1553-1560XuL, ZhuL F, ZhangX L. Research on resistance mechanism of cotton to Verticillium wilt. Acta Agron Sin, 2012, 38: 1553-1560[本文引用:1][CJCR: 1.667]
[20]
YangC, GuoW Z, LiG, GaoF, LinS, ZhangT. QTL mapping for Verticillium wilt resistance at seedling and maturity stages in Gossypium barbadense L. Plant Sci, 2008, 174: 290-298[本文引用:1][JCR: 2.922]
[21]
RodriguezM C, PetersenM, MundyJ. Mitogen-activated protein kinase signaling in plants. Annu Rev Plant Biol, 2010, 61: 621-649[本文引用:1][JCR: 25.962]
[22]
delPozo O, PedleyK F, MartinG B. MAPKKKa is a positive regulator of cell death associated with both plant immunity and disease. EMBO J, 2004, 23: 3072-3082[本文引用:1][JCR: 9.822]
[23]
JonesL, RztcliffF, BaulcombeD C. RNA-induced transcriptional gene silencing in plants can be inherited independently of the RNA trigger and requires Met1 for maintenance. Curr Biol, 2001, 11: 747-757[本文引用:1][JCR: 9.494]
[24]
LiuE W, PageJ E. Optimized cDNA libraries for virus-induced gene silencing (VIGS) using tobacco rattle virus. Plant Methods, 2008, 4: 5[本文引用:1][JCR: 2.667]
[25]
PeeleC, JordanC V, MuangsanN, TurnageM, EgelkroutE, EagleP, Hanley-BowdoinL, RobertsonD. Silencing of a meristematic gene using geminivirus-derived vectors. Plant J, 2001, 27: 357-366[本文引用:1][JCR: 6.582]
[26]
VoinnetO, LedererC, BaulcombeD C. A viral movement protein prevents spread of the gene silencing signal in Nicotiana benthamiana. Cell, 2000, 103: 157[本文引用:1][JCR: 31.957]
[27]
WatsonJ M, WangM B. Antiviral resistance in plants: methods and protocols. New York: Humana Press, 2012, p894, DOIDOI:10.1007/978-1-61779-882-5_6[本文引用:1]
[28]
GuoW, CaiC, WangC, ZhaoL, WangL, ZhangT. A preliminary analysis of genome structure and composition in Gossypium hirsutum. BMC Genomics, 2008, 9: 314[本文引用:1][JCR: 4.397]
[29]
GaoW, LongL, ZhuL F, XuL, GaoW H, SunL Q, LiuL L, ZhangX L. Proteomic and virus-induced gene silencing (VIGS) analyses reveal that gossypol, brassinosteroids, and jasmonic acid contribute to the resistance of cotton to Verticillium dahliae. Mol Cell Proteom, 2013, 12: 3690-3703[本文引用:1][JCR: 7.251]
 , 吕坤
, 吕坤









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}