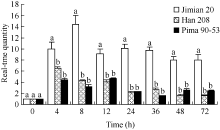
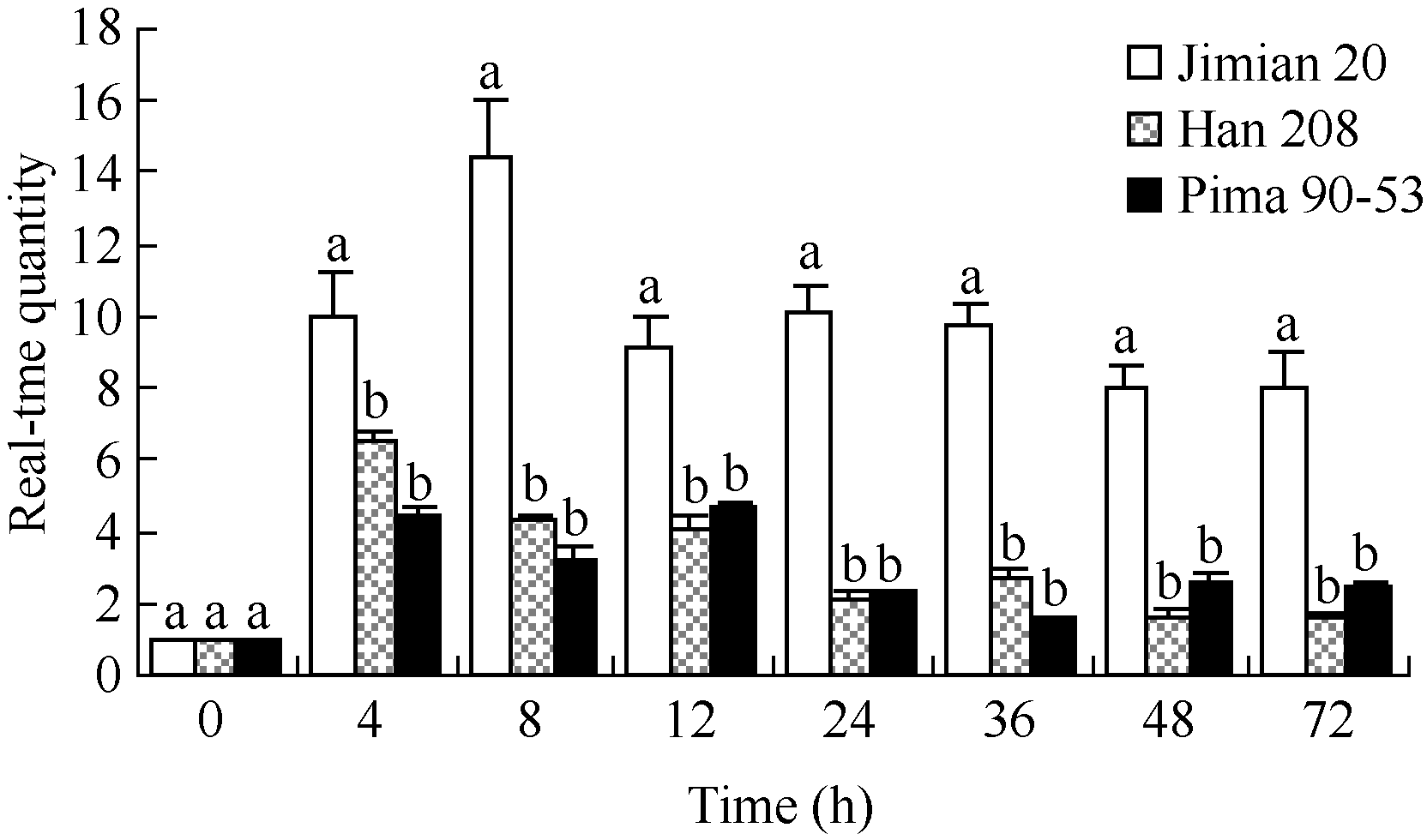
关键词:棉花; 黄萎病; 木质素; 漆酶 Function of Acid Insoluble Lignin andGhLaccase in Cotton Resistance to Verticillium wilt WU Li-Zhu, WANG Xing-Fen, ZHANG Yan, LI Xi-Huan, ZHANG Gui-Yin, WU Li-Qiang, LI Zhi-Kun, MA Zhi-Ying* North China Key Laboratory for Germplasm Resources of Education Ministry / Key Laboratory of Crop Germplasm Resources of Hebei / Agricultural University of Hebei, Baoding 071001, China Fund: AbstractTo search cotton resistance to Verticillium wilt, we inoculatedVerticillium dahliae into roots of three cotton varieties with different resistance, Pima 90-53, Jimian 20, and Han 208, observed the structure of cell wall, determined the content of lignin and analyzed the transcript level ofLaccase. The results showed that, at one day after infection (dpi), the pathogen was unobserved in root vascular tissue of Pima 90-53, while observed in Jimian 20 and Han 208. At 35 days after infection, the cell wall was highly lignified and the vascular was unblocked in Pima 90-53, while there were moderately lignified and slightly blocked in Jimian 20, and lowly lignified and seriously blocked in Han 208. The contents of acid soluble lignin (ASL) and acid insoluble lignin (AIL) in leaves and petioles of the three cotton varieties planted in the diseased nursery were measured. The results showed that the content of AIL was negatively correlated with the disease index of the three cultivars (r= 0.99991*), and the cotent ratio of AIL to ASL was in accord with the verticillium wilt resistance of cotton. The transcript level ofLaccase was detected using real-time PCR method, the transcription level ofGhLaccase in Jimian 20 was significantly higher than that in Han 208 at any point of test time. The expression level reached the highest at 8 h after infection and maintained the high level within three days. SoGhLaccase performed high efficiency in response to the infection ofVerticillium dahliae. In conclusion, AIL is positively correlated with the verticillium wilt resistance of cotton, andLaccase plays an important role in cotton resistance toVerticillium dahliae.
表1 病圃中不同抗性棉花品种细胞壁木质素含量 Table 1 Content of lignin in three different cotton varieties in the disease nursery
成分 Component
组织 Organ
邯208 Han 208
冀棉20 Jimian 20
比马90-53 Pima 90-53
酸可溶性木质素 Acid soluble lignin (ASL)
叶片Leaf
1.58±0.02 A
1.54±0.02 A
1.31±0.01 B
叶柄Petiole
1.93±0.01 a
1.81±0.00 b
1.33±0.04 c
酸不可溶性木质素 Acid insoluble lignin (AIL)
叶片Leaf
12.22±2.60 c
14.51±1.42 b
17.07±0.59 a
叶柄Petiole
14.90±1.69 b
19.30±1.27 a
20.36±1.09 a
酸不可溶性木质素/酸可溶性木质素 AIL/ASL
叶片Leaf
7.71±1.55 b
9.42±0.78 b
13.05±0.26 a
叶柄Petiole
7.71±0.80 c
10.66±0.65 b
15.35±0.31 a
标以不同大小写字母值分别表示在0.01或0.05的水平上差异显著。 Values followed by different capital and small letters are significantly different at 0.01 or 0.05 probability levels, respectively.
表1 病圃中不同抗性棉花品种细胞壁木质素含量 Table 1 Content of lignin in three different cotton varieties in the disease nursery
马存, 陈其煐. 我国棉花抗枯、黄萎病育种研究进展. 中国农业科学, 1992, 25(1): 50-57MaC, ChenQ Y. Progress on studies of cotton resistant breeding fusarium and verticillium wilts in China. Sci Agric Sin, 1992, 25(1): 50-57 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[2]
简桂良, 邹亚飞, 马存. 棉花黄萎病连年流行的原因及对策. 中国棉花, 2003, 30(3): 13-14JianG L, ZouY F, MaC. Current status and countermeasure of verticillium wilt of cotton in China. China Cotton, 2003, 30(3): 13-14 (in Chinese)[本文引用:1][CJCR: 0.3221]
[3]
KotchoniS O, GachomoE W. The reactive oxygen species network pathways: an essential prerequisite for perception of pathogen attack and the acquired disease resistance in plants. J Biosci, 2006, 31: 389-404[本文引用:1][JCR: 1.759]
[4]
陈捷胤, 戴小枫. 棉花对黄萎病的抗病机制研究进展. 分子植物育种, 2005, 3: 427-435ChenJ Y, DaiX F. Research advance on the resistant mechanism of cotton against Verticillium wilt. Mol Plant Breed, 2005, 3: 427-435 (in Chinese with English abstract)[本文引用:1][CJCR: 1.23]
[5]
SmitF, DuberyI A. cell wall reinforcement in cotton hypocotyls inresponse to a Verticillium dahliae elicitor. Phytochemistry, 1997, 44: 811-815[本文引用:4][JCR: 3.05]
[6]
DongH Z, LiW J, ZhangD M, TangW. Differential expression of induced resistance by an aqueous extract of killed Penicillium chrysogenum against verticillium wilt of cotton. Crop Prot, 2003, 22: 129-134[本文引用:1][JCR: 1.303]
[7]
王莉. 棉株受黄萎病菌侵染后组织结构变化与抗性的相互关系. 农业技术与装备, 2012, 22(11): 32-34WangL. The relationgshap between resistentant and the tissue construction of cotton indefectted by Verticillium dahliae. Agric Technol Equip, 2012, 22(11): 32-34 (in Chinese)[本文引用:1]
[8]
侯丽娟, 李卫, 刘燕霞, 杨家荣. 棉花黄萎病菌毒素对棉花生化代谢的影响. 西北农业学报, 2010, 19(12): 63-67HouL J, LiW, LiuY X, YangJ R. Effect of V. dahliae toxin on biochemical metabolism of cotton seedlings. Acta Agric Borea-li-Occident Sin, 2010, 19(12): 63-67 (in Chinese with English abstract)[本文引用:1]
[9]
HosokawaM, SuzukiS, UmezawaT, SatoY. Progress of lignification mediated by intercellular transportation of monolignols during tracheary element differentiation of isolated Zinnia mesophyll cells. Plant Cell Physiol, 2001, 42: 959-968[本文引用:1][JCR: 4.134]
[10]
Bland D E, SkickoJ, MenshunM. The relationship of acid insoluble lignin and acid soluble lignin to the lignins of the middle lamella and cell wall in Eucalyptus regnans. Holzforschung, 1975, 29: 144-147[本文引用:1][JCR: 2.416]
[11]
XuL, ZhuL F, TuL L, LiuL L, YuanD J, JinL, LongL, ZhangX L. Lignin metabolism has a central role in the resistance of cotton to the wilt fungus Verticillium dahliae as revealed by RNA-Seq-dependent transcriptional analysis and histochemistry. J Exp Bot, 2011, 62: 5607-5621[本文引用:3][JCR: 5.242]
[12]
ZhangY, WangX F, YangS, ChiJ N, ZhangG Y, MaZ Y. Cloning and characterization of a verticillium wilt resistance gene from Gossypium barbadense and functional analysis in Arabidopsis thaliana. Plant Cell Rep, 2011, 30: 2085-2096[本文引用:1][JCR: 2.509]
王省芬, 田海燕, 马峙英, 张桂寅, 玄兆伶, 王文生, 孙艳香. 黄萎病菌诱导下陆地棉抗病品种SSH文库的构建. 棉花学报, 2008, 20: 3-8WangX F, TianH Y, MaZ Y, ZhangG Y, XuanZ L, WangW S, SunY X. SSH library construction of upland cotton resistant cultivar under the stress of Verticillium dahliae. Cotton Sci, 2008, 20: 3-8 (in Chinese with English abstract)[本文引用:2][CJCR: 1.462]
[15]
张纯颖, 王省芬, 张桂寅, 吴立强, 迟吉娜, 李志坤, 马峙英. 黄萎病菌诱导下陆地棉抗病品种SSH文库的EST分析. 棉花学报, 2010, 22: 17-22ZhangC Y, WangX F, ZhangG Y, WuL Q, ChiJ N, LiZ K, MaZ Y. ESTs analysis of suppression subtractive hybridization library from upland cotton resistant cultivar infected by Verticillium dahliae. Cotton Sci, 2010, 22: 17-22 (in Chinese with English abstract)[本文引用:2][CJCR: 1.462]
[16]
LivakK J, SchmittgenT D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25: 402-408[本文引用:1][JCR: 3.641]
[17]
张桂寅, 吴立强, 李志坤, 工省芬, 马峙英. 不同抗性品种对棉花黄萎病菌致病力的影响. 棉花学报, 2012, 24: 529-534ZhangG Y, WuL Q, LiZ K, WangX F, MaZ Y. Effect of host resistance on the pathogenicity of Verticillium dahliae isolates. Cotton Sci, 2012, 24: 529-534 (in Chinese with English abstract)[本文引用:2][CJCR: 1.462]
[18]
ZhangY, WangX F, DingZ G, MaQ, ZhangG R, ZhangS L, LiZ K, WuL Q, ZhangG Y, MaZ Y. Transcriptome profiling of Gossypium barbadense inoculated with Verticillium dahliae provides a resource for cotton improvement. BMC Genomic, 2013, 14: 637-655[本文引用:1][JCR: 4.397]
[19]
HuismanO C. Interrelations of root growth dynamics to epidemiology of root-invading fungi. Ann Rev Phytopathol, 1982, 20: 303-327[本文引用:1]
[20]
DebodeJ, ClewesE, BackerD D, HofteaM. Lignin is involved in the reduction of Verticillium dahliae var. longisporum inoculum in soil by crop residue incorporation. Soil Biol Biochem, 2005, 37: 301-309[本文引用:1][JCR: 3.654]
张松贺, 马荣才. 伤害和软腐病菌侵染引起大白菜木质素合成途径基因表达水平提高及木质素含量增加. 中国农业生物技术学会第三届会员代表大会暨学术交流会论文摘要集, 2006. p64ZhangS H, MaR C. The Content of Lignin Improved in Chinese Cabbage and the Trascription Level of Genes in the Lignin Synthesis Pathway Increased Induced by Damage and Erwinia carotovora. Proceedings of the Third China Agricultural Biotechno-logy Academic Communication and Representative Assembly, 2006. p64 (in Chinese)[本文引用:1]
BaoW, O’MalleyD M, WhettenR, SederoffR R. A laccase associated with lignification in loblolly pine xylem. Science, 1993, 260: 672-674[本文引用:1]
[25]
BerthetS, Demont-CauletN, PolletB, BidzinskiP, CezardL, BrisP L, BorregaN, HerveJ, BlondetE, BalzergueS, LapierreC, JouaninL. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant Cell, 2011, 23: 1124-1137[本文引用:1][JCR: 9.251]
[26]
DeanJ F D, LaFayetteP R, RughC, TristramA M, HoopesJ T, ErikssonK E L, MarkleS A. Laccase associated with lignifying vascular tissues. In: Lewis N G, Sarkanen S, eds. Lignin and lignan biosynthesis. American Chemical Society, Washington, DC. 1998. pp96-108[本文引用:1]
[27]
LiL, SteffensJ C. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta, 2002, 215: 239-247[本文引用:1][JCR: 3.347]
[28]
王骥, 朱木兰, 卫志明. 棉花漆酶基因在转基因新疆杨中的表达及其对木质素合成的影响. 分子细胞生物学报, 2008, 41: 11-18WangJ, ZhuM L, WeiZ M. Cotton laccase gene overexpression in transgenic Populus alba var. pyramidalis and its effects on the lignin biosynthesis in transgenic plants. J Mol Cell Biol, 2008, 41: 11-18 (in Chinese with English abstract)[本文引用:1][CJCR: 0.487]
[29]
WangG D, LiQ J, LuoB, ChenX Y. Ex planta phytoremediation of trichlorophenol and phenolic allelochemicals via an engineered secretory Laccasecase. Nat Biotechnol, 2004, 22: 893-897[本文引用:1][JCR: 32.438]
 , 王省芬, 张艳, 李喜焕, 张桂寅, 吴立强, 李志坤, 马峙英
, 王省芬, 张艳, 李喜焕, 张桂寅, 吴立强, 李志坤, 马峙英






{kind=link}
{kind=link}
{kind=link}
{kind=link}