关键词:棉花; 甲哌鎓; 柯巴基焦磷酸合酶; 内根-贝壳杉烯合酶; 叶面积 GhCPS andGhKS Encoding Gibberellin Biosynthesis Enzymes Involve in Inhibition of Leaf Growth by Mepiquat Chloride in Cotton (Gossypium hirsutum L.) WANG Li1,2, ZHANG Ming-Cai1, DU Ming-Wei1, TIAN Xiao-Li1, LI Zhao-Hu1,* 1Engineering Research Center of Plant Growth Regulator, Ministry of Education / College of Agronomy and Biotechnology, China Agricultural University, Beijing 10093, China
2College of Life Science, Henan Normal University, Xinxiang 453007, China
Fund: AbstractEnt-copalyl diphosphate synthase (CPS) and ent-kaurene synthase (KS) are the key enzymes involved in the early steps of gibberellin (GA) biosynthesis. This paper aimed at elucidating whether the action of mepiquat chloride (DPC) on leaf growth was related to the expression levels ofGhCPSandGhKS in cotton seedlings. DPC was foliar applied to seedlings at the 3rd leaf expanded stage of cotton cultivar Xinkang 4 by pot culture. The results showed that DPC significantly decreased the leaf area, and the area of the 4th leaf was decreased more than that of the 3rd leaf. DPC at 80 mg L-1 markedly reduced GA4 content in the 3rd leaf at four days after treatment and in the 4th leaf from four to six days after treatment. The expression levels ofGhCPS andGhKS in the 3rd leaf were decreased by DPC from one to four days after treatment, and similar trends were observed in the 4th leaf from one to six days after treatment. All the results suggested that DPC could reduce endogenous GA4 content by downregulatingGhCPS andGhKS expressions, leading to a smaller leaf size. Otherwise, the younger leaf was more sensitive to DPC.
Keyword:Cotton; Mepiquat chloride; Ent-copalyl diphosphate synthase; Ent-kaurene synthase; Leaf area Show Figures Show Figures
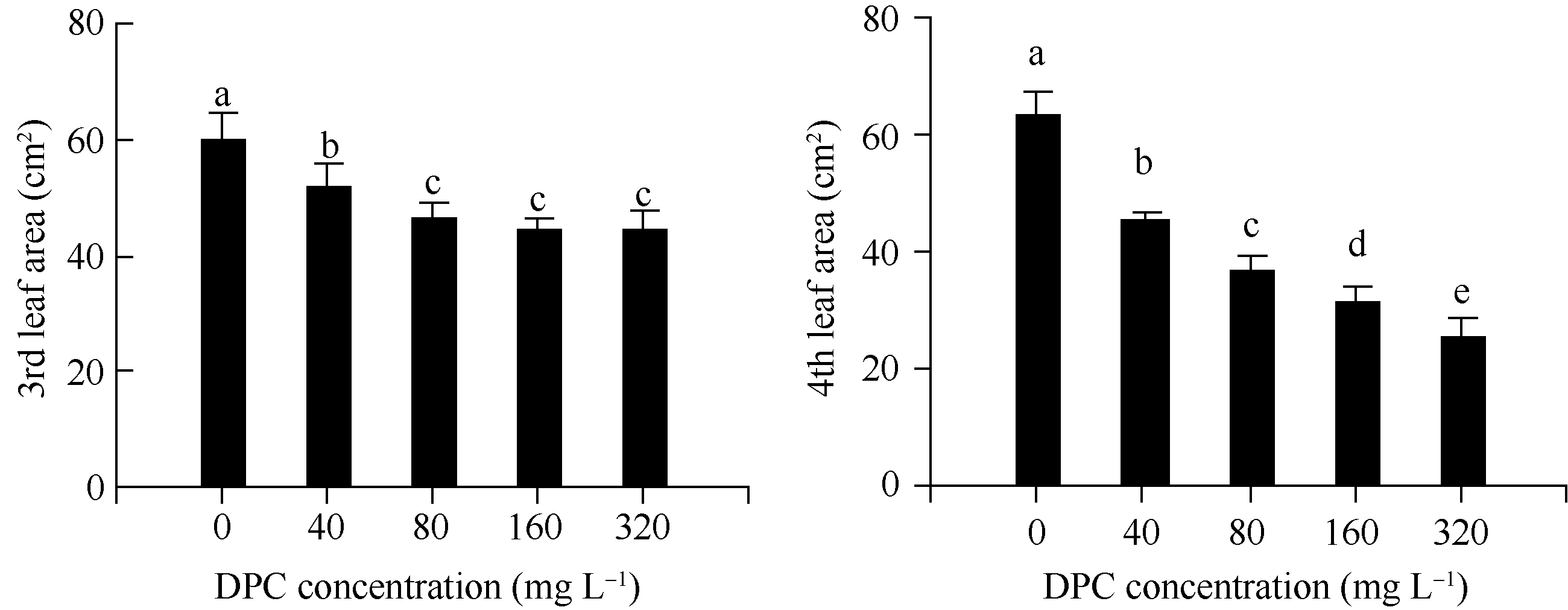
图1 不同浓度DPC对棉花幼苗第3和第4叶叶面积的影响DPC: 甲哌鎓; 数据为平均值± SE; 标以不同字母的柱值在0.05水平上差异显著。Fig. 1 Effect of different concentrations of mepiquat chloride on the 3rd and 4th leaf area in cotton seedlingsDPC: mepiquat chloride; data are means± SE; Bars superscripted by different letters are significantly different ( P<0.05).
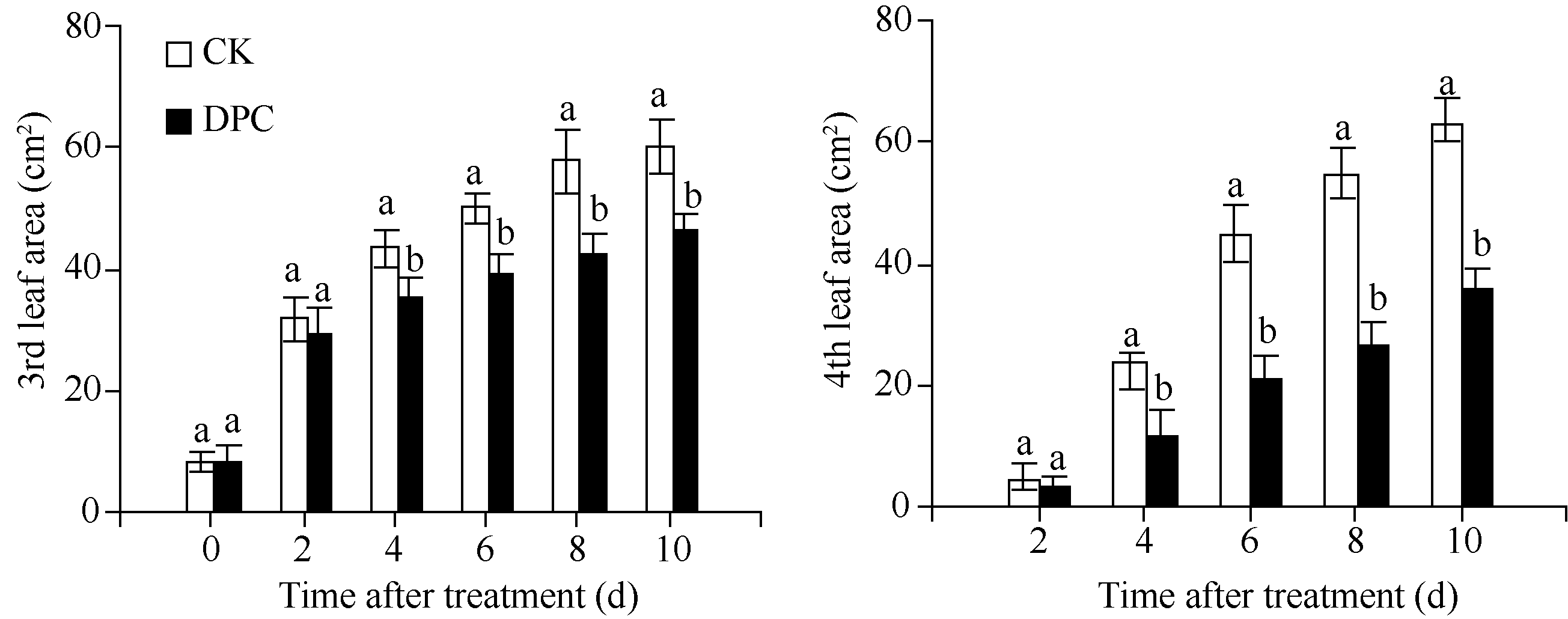
图2 DPC对棉花幼苗第3和第4叶叶面积扩展动态的调控DPC: 80 mg L-1甲哌鎓; 数据为平均值± SE; 标以不同字母的柱值在0.05水平上差异显著。Fig. 2 Regulation of mepiquat chloride on the expansion of 3rd and 4th leaf area in cotton seedlingsDPC: 80 mg L-1 mepiquat chloride; data are means± SE; Bars superscripted by different letters are significantly different ( P<0.05).
图3 DPC对棉花幼苗第3和第4叶GA4含量的动态调控DPC: 80 mgL-1甲哌鎓; 数据为平均值±SE; 标以不同字母的柱值在0.05水平上差异显著。Fig. 3 Regulation of mepiquat chloride on GA4 content in the 3rd and 4th leaves of cotton seedlingsDPC: 80 mg L-1 mepiquat chloride; data are means± SE; Bars superscripted by different letters are significantly different ( P<0.05).
图4 DPC对棉花幼苗第3和第4叶 GhCPS和 GhKS表达的动态调控DPC: 80 mg L-1甲哌鎓; 数据为平均值± SE; 标以不同字母的柱值在0.05水平上差异显著。Fig. 4 Regulation of mepiquat chloride on expressions of GhCPS and GhKS in the 3rd and 4th leaves in cotton seedlingsDPC: 80 mg L-1 mepiquat chloride; data are means± SE; Bars superscripted by different letters are significantly different ( P<0.05).
SiebertJ D, StewartA M. Influence of plant density on cotton response to mepiquat chloride application. Agron J, 2006, 98: 1634-1639[本文引用:2][JCR: 1.518]
[2]
RenX, ZhangL, DuM, EverscJ B, WerfW, TianX, LiZ. Managing mepiquat chloride and plant density for optimal yield and quality of cotton. Field Crops Res, 2013, 149: 1-10[本文引用:2][JCR: 2.474]
[3]
ReddyV R, BakerD N, HodgesH F. Temperature and mepiquat chloride effects on cotton canopy architecture. Agron J, 1990, 82: 190-195[本文引用:3][JCR: 1.518]
[4]
ReddyA R, ReddyK R, HodgesH F. Mepiquat chloride (PIX) induced changes in photosynthesis and growth of cotton. Plant Growth Regul, 1996, 20: 179-183[本文引用:3][JCR: 1.67]
[5]
ZhaoD, OosterhuisD M. Pix plus and mepiquat chloride effects on physiology, growth, and yield of field-grown cotton. J Plant Growth Regul, 2000, 19: 415-422[本文引用:1][JCR: 1.99]
[6]
GoniasE D, OosterhuisD M, BibiA C. Cotton radiation use efficiency response to plant growth regulators. J Agric Sci, 2012, 150: 595-602[本文引用:2][JCR: 2.041]
[7]
RademacherW. Growth retardants: effects on gibberellin biosynthesis and other metabolic pathways. Annu Rev Plant Physiol Mol Biol, 2000, 51: 501-531[本文引用:2][JCR: 25.962]
[8]
DennisD T, UpperC D, WestC A. An enzymic site of inhibition of gibberellin biosynthesis by Amo 1618 and other plant growth retardants. Plant Physiol, 1965, 40: 948-952[本文引用:1][JCR: 6.555]
[9]
ShechterI, WestC A. Biosynthesis of Gibberellins. IV. Biosynthesis of cyclic diterpenes from trans-geranylgeranyl pyrophosphate. J Biol Chem, 1969, 244: 3200-3209[本文引用:1][JCR: 4.651]
[10]
SmithM W, YamaguchiS, Ait-AliT, KamiyaY. The first step of gibberellin biosynthesis in pumpkin is catalyzed by at least two copalyl diphosphate synthases encoded by differentially regulated genes. Plant Physiol, 1998, 118: 1411-1419[本文引用:3][JCR: 6.555]
[11]
SilverstoneA L, ChangC, KrolE, SunT P. Developmental regulation of the gibberellin biosynthetic gene GA1 in Arabidopsis thaliana. Plant J, 1997, 12: 9-19[本文引用:2][JCR: 6.582]
[12]
KoornnefM, van der Veen J H. Induction and analysis of gibberellin-sensitive mutants in Arabidopsis thaliana (L. ) Heynh. Theor Appl Genet, 1980, 58: 257-263[本文引用:5][JCR: 3.658]
[13]
SunT, KamiyaY. The Arabidopsis GAl locus encodes the cyclase ent-kaurene synthetase A of gibberellin biosynthesis. Plant Cell, 1994, 6: 1509-1518[本文引用:5][JCR: 9.251]
何钟佩. 农作物化学控制实验指导. 北京: 北京农业大学出版社, 1993. pp36-39HeZ P. Experimental guide of chemical control of crops. Beijing: Beijing Agricultural University Press, 1993. pp36-39(in Chinese)[本文引用:1]
[16]
LivakK J, SchmittgenT D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods, 2001, 25: 402-408[本文引用:1][JCR: 3.641]
[17]
BarbosaL M, Castro P R C. Comparison between concentrations and application time of mepiquat chloride, chlorocholine chloride and ethephon in cotton (Gossypium hirsutum L. cv. IAC-17). Planta Daninha, 1983, 6: 1-10[本文引用:1][JCR: 1.204]
[18]
FernándezC J, CothrenJ T, McInnesK J. Partitioning of biomass in well-watered and water-stressed cotton plants treated with mepiquat chloride. Crop Sci, 1991, 31: 1224-1228[本文引用:1][JCR: 1.513]
OlszewskiN, SunT P, GublerF. Gibberellin signaling: biosynthesis, catabolism, and response pathways. Plant Cell, 2002, 14: 61-80[本文引用:1][JCR: 9.251]
[21]
YamaguchiS. Gibberellin metabolism and its regulation. Annu Rev Plant Biol, 2008, 59: 225-251[本文引用:1][JCR: 25.962]
[22]
JiangX, LiH, WangT, PengC, WangH, WuH, WangX. Gibberellin indirectly promotes chloroplast biogenesis as a means to maintain the chloroplast population of expand ed cells. Plant J, 2012, 72: 768-780[本文引用:1][JCR: 6.582]
KangS M, KimbJ T, HamayunM, HwangI C, KhanA L, KimY H, LeeJ H, LeeI J. Influence of prohexadione-calcium on growth and gibberellins content of Chinese cabbage grown in alpine region of South Korea. Sci Hortic, 2010, 125: 88-92[本文引用:1][JCR: 1.396]
[25]
OtaniM, MeguroS, GondairaH, HayashiM, SaitoM, HanD S, InthimaP, SupaibulwatanaK, MoriS, JikumaruY, KamiyaY, LiT, NikiT, NishijimaT, KoshiokaM, NakanoM. Overexpression of the gibberellin 2-oxidase gene from Torenia fournieri induces dwarf phenotypes in the liliaceous monocotyledon Tricyrtis sp. J Plant Physiol, 2013, 170: 1416-1423[本文引用:1][JCR: 2.699]
[26]
YamaguchiS, SunT P, KawaideH, KamiyaY. The GA2 locus of Arabidopsis thaliana encodes ent-kaurene synthase of gibberellin biosynthesis. Plant Physiol, 1998, 116: 1271-1278[本文引用:1]
[27]
AyeleB T, OzgaJ A, KurepinL V, ReineckeD M. Developmental and embryo axis regulation of gibberellin biosynthesis during germination and young seedling growth of pea. Plant Physiol, 2006, 142: 1267-1281[本文引用:1][JCR: 6.555]
[28]
YamaguchiS, KamiyaY, SunT P. Distinct cell-specific expression patterns of early and late gibberellin biosynthetic genes during Arabidopsis seed germination. Plant J, 2001, 28: 443-453[本文引用:1][JCR: 6.582]
[29]
李晨晨, 侯雷, 尹亮, 赵金凤, 袁守江, 张文会, 李学勇. 水稻极矮突变体s2-47对赤霉素的响应及基因定位研究. 作物学报, 2013, 39: 1766-1774LiC C, HouL, YinL, ZhaoJ F, YuanS J, ZhangW H, LiX Y. Gibberellin responsiveness and gene mapping of rice extreme dwarf mutant s2-47. Acta Agron Sin, 2013, 39: 1766-1774 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]








{kind=link}
{kind=link}
{kind=link}
{kind=link}