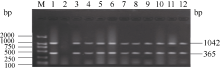
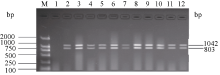
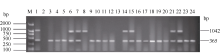
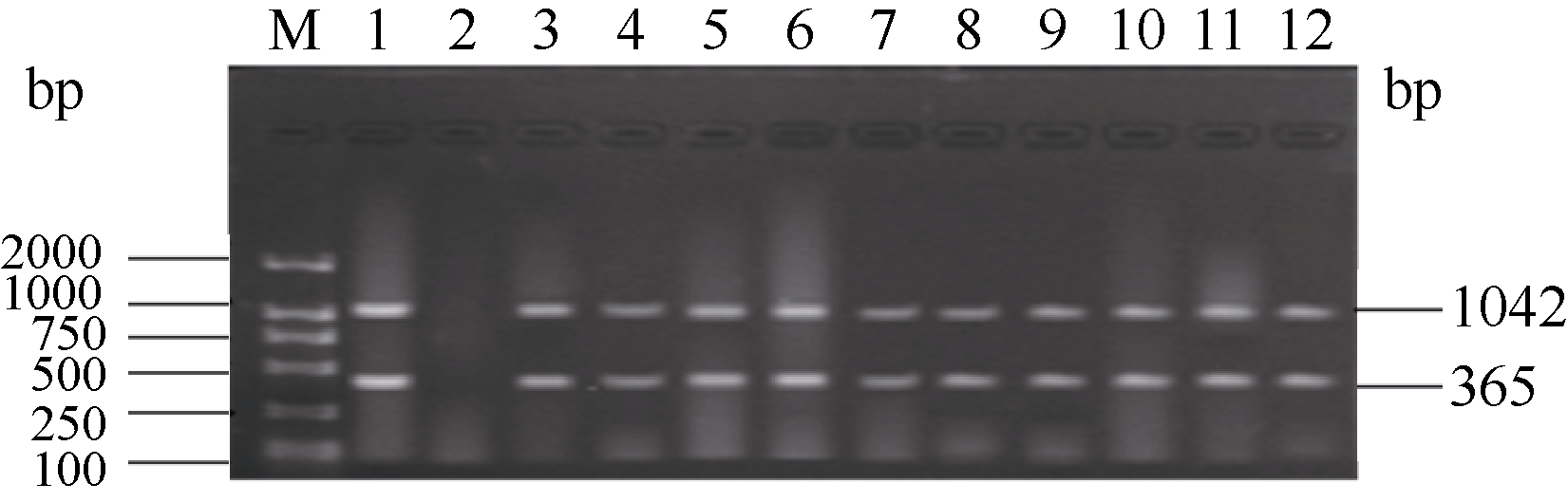
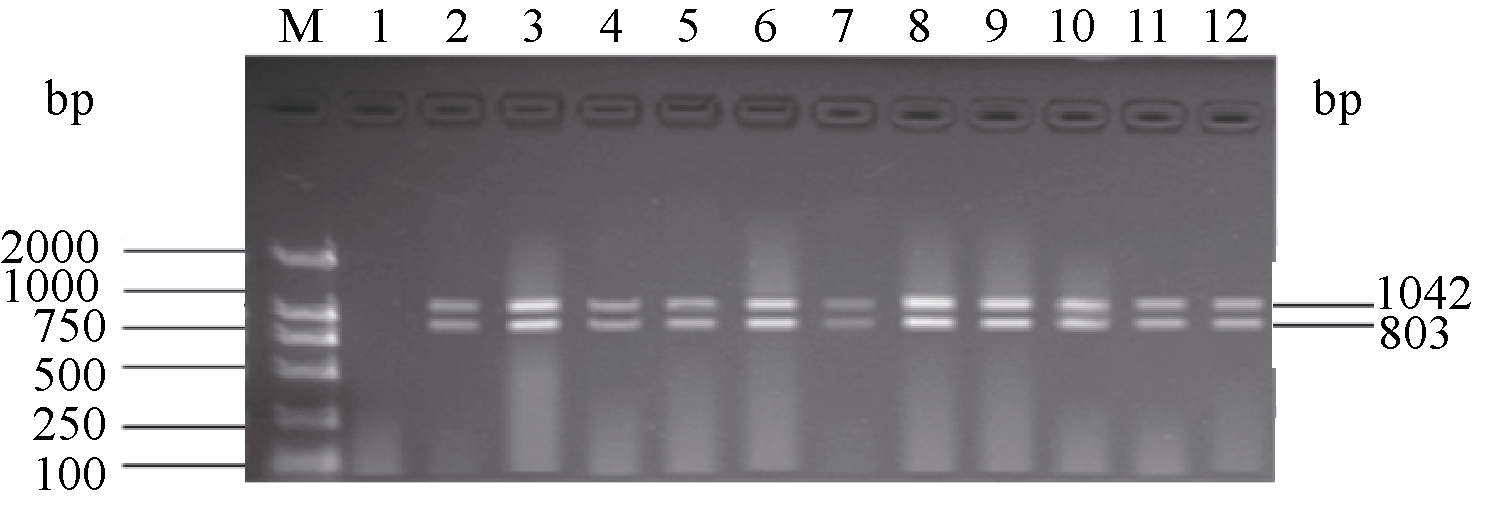
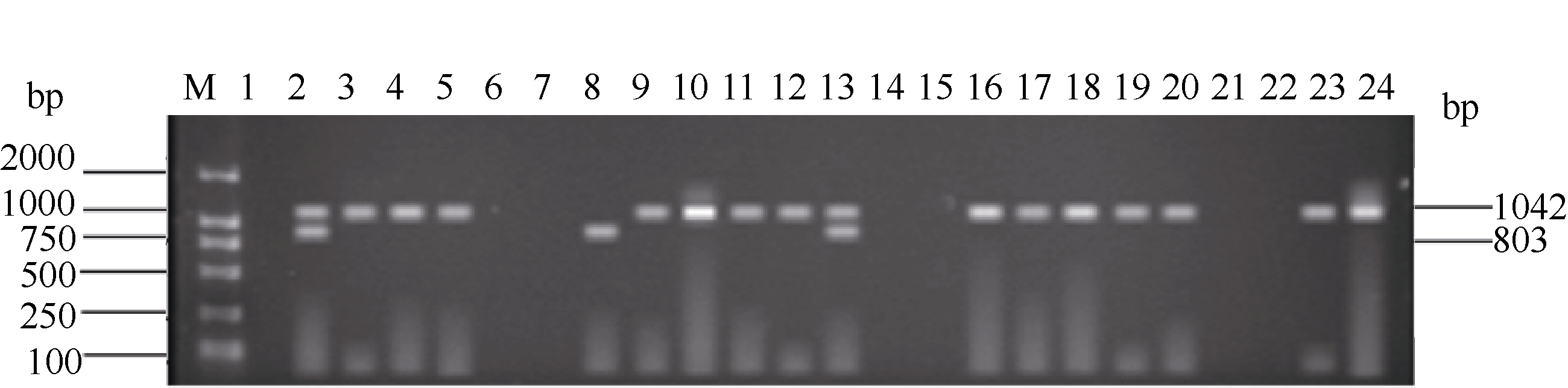
关键词:稻瘟病; 分子标记; 抗性基因; 多重PCR体系 Establishment and Application of Multiplex PCR System for Blast Resistance GenesPi-ta andPi-bin Rice YAO Shu, LIU Yan-Qing, ZHANG Ya-Dong, ZHU Zhen, CHEN Tao, ZHAO Qing-Yong, ZHOU Li-Hui, ZHAO Chun-Fang, YU Xin, WANG Cai-Lin* Institute of Food Crops, Jiangsu Academy of Agricultural Sciences / Jiangsu High Quality Rice R&D Center / Nanjing Branch of China National Center for Rice Improvement, Nanjing 210014, China Fund: AbstractRice blast is one of the important diseases in major rice producing areas in China. The main blast resistance genesPi-taandPi-b showed wide and durable resistance spectrum in many rice growing areas. In this study, two sets of multiple PCR system primed by functional markers were established to identifyPi-ta andPi-b, using 22 rice varieties with known alleles on bothPi-ta andPi-bloci. The PCR system I could detect the resistance alleles onPi-ta andPi-b loci, while the PCR system II could detect the susceptible alleles simultaneously. The effectiveness of the two PCR systems was validated in 336japonica breeding lines. The result was highly consistent with that of molecular detection using conventional single marker. Therefore, the two multiple PCR systems developed are stable, reliable and time-saving, and can serve as a rapid and efficient method to identifyPi-taandPi-b genes in bulky screening and marker-assistant selection in rice aiming at blast resistance breeding.
Keyword:Rice blast; Molecular marker; Resistance gene; Multiplex PCR system Show Figures Show Figures
郑钊, 陈由强, 张建福, 谢华安. 水稻稻瘟病抗性基因的定位、克隆及应用. 分子植物育种, 2009, 7: 385-392ZhengZ, ChenY Q, ZhangJ F, XieH A. Mapping cloning of rice blast resistance genes and their application. Mol Plant Breed, 2009, 7: 385-392 (in Chinese with English abstract)[本文引用:1][CJCR: 1.23]
[2]
沈瑛, 朱培良, 袁筱萍, LevyM, DeckerM, TalbotN, HamerJ E. 中国稻瘟病菌的遗传多样性. 植物病理学报, 1993, 23: 309-313ShenY, ZhuP L, YuanX P, LevyM, DeckerM, TalbotN, HamerJ E. Genetic diversity of Magnaporthe grisea in China. Acta Phytopathol Sin, 1993, 23: 309-313 (in Chinese)[本文引用:1][CJCR: 1.088]
[3]
HittalmaniS, ParcoA, MewT V, ZeiglerR S, HuangN. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor Appl Genet, 2000, 100: 1121-1128[本文引用:1][JCR: 3.658]
[4]
ZhaiC, LinF, DongZ Q, HeX Y, YuanB, ZengX S, WangL, PanQ H. The isolation and characterization of Pik, a rice blast resistance gene which emerged after rice domestication. New Phytol, 2011, 189: 321-334[本文引用:1][JCR: 6.736]
[5]
HayashiN, InoueH, KatoT, ShirotaM, KanamoriH, YamaneH, Hayano-SaitoY, MatsumotoT, YanoM, TakatsujiH. Durable panicle blast resistance gene Pb1 encodes an atypical CC-NBS- LRR protein and was generated by acquiring a promoter through local genome duplication. Plant J, 2010, 64: 498-510[本文引用:1][JCR: 6.582]
[6]
OkuyamaY, KanzakiH, AbeA, YoshidaK, TamiruM, SaitohH, FujibeT, MatsumuraH, ShentonM, GalamD C, UndanJ, ItoA, SoneT, TerauchiR. A multifaceted genomics approach allows the isolation of the rice Pia-blast resistance gene consisting of two adjacent NBS-LRR protein genes. Plant J, 2011, 66: 467-479[本文引用:1][JCR: 6.582]
[7]
YuanB, ZhaiC, WangW J, ZengX S, XuX K, HuH Q, LinF, WangL, PanQ H. The Pik-p resistance to Magnaporthe oryzae in rice is mediated by a pair of closely linked CC-NBS-LRR genes. Theor Appl Genet, 20l1, 122: 1017-1028[本文引用:1]
[8]
TakahashiA, HayashiN, MiyaoA, HirochikeH. Unique features of the rice blast resistance Pish locus revealed by large scale ret retrotransposon-tagging. BMC Plant Biol, 2010, 10: 175[本文引用:1][JCR: 4.354]
[9]
WangZ X, YanoM, YamanouchiU, IwamotoM, MonnaL, HayasakaH, KatayoseY, SasakiT. The Pi-b gene for rice blast resistance belongs to the nucleotide binding and leucine rich repeat class of plant resistance genes. Plant J, 1999, 19: 55-64[本文引用:1][JCR: 6.582]
[10]
BryanG T, WuK S, FarrallL, JiaY L, HersheyH P, McadamsS A, FaultK N, DonaldsonG K, RenatoT, ValentB. A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance gene Pi-ta. Plant Cell, 2000, 12: 2033-2045[本文引用:1][JCR: 9.251]
[11]
WangZ, JiaY, RutgerJ N, XiaY, Rapid survey for presence of a blast resistance gene Pi-ta in rice cultivars using the dominant DNA markers derived from portions of the Pi-ta gene. Plant Breed, 2007, 126: 36-42[本文引用:1][JCR: 1.175]
[12]
刘洋, 徐培洲, 张红宇, 徐建第, 吴发强, 吴先军. 水稻抗稻瘟病Pi-b基因的分子标记辅助选择与应用. 中国农业科学, 2008, 41: 9-14LiuY, XuP Z, ZhangH Y, XuJ D, WuF Q, WuX J. Marker-assisted selection and application of blast resistant gene Pib in rice. Sci Agric Sin, 2008, 41: 9-14 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[13]
李进斌, 王甜, 许明辉. 云南地方稻种抗稻瘟病基因Pi-ta和Pi-b的鉴定. 中国水稻科学, 2012, 26: 593-599LiJ B, WangT, XuM H. Identification of Pi-ta and Pi-b genes for rice blast resistance of rice land races from Yunnan Province. Chin J Rice Sci, 2012, 26: 593-599 (in Chinese with English abstract)[本文引用:2][CJCR: 1.494]
[14]
刘华招, 刘延, 刘化龙, 徐正进, 陈温福. 黑龙江省种植品种中稻瘟病抗性基因Pi-b和Pi-ta的分布. 东北农业大学学报, 2011, 42(1): 27-31LiuH Z, LiuY, LiuH L, XuZ J, ChenW F. Distribution of two blast resistance genes Pib and Pita in major rice cu1tivars in Heilongjiang Province in China. J. Northeast Agric Univ, 2011, 42(1): 27-31 (in Chinese with English abstract)[本文引用:1][CJCR: 0.1053]
[15]
时克, 雷财林, 程治军, 许兴涛, 王久林, 万建民. 稻瘟病抗性基因Pi-ta和Pi-b在我国水稻主栽品种中的分布. 植物遗传资源学报, 2009, 10: 21-26ShiK, LeiC L, ChengZ J, XuX T, WangJ L, WanJ M. Distribution of two blast resistance genes Pita and Pi-b in major rice cultivars in China. J Plant Genet Resour, 2009, 10: 21-26 (in Chinese with English abstract)[本文引用:2][CJCR: 1.1628]
[16]
MaW, ZhangW, GaleK R. Multiplex-PCR typing of high molecular weight glutenin alleles in wheat. Euphytica, 2003, 134: 51-60[本文引用:1][JCR: 1.643]
[17]
NakamuraT, VrintenP, SaitoM, KondaM. Rapid classification of partial waxy wheat using PCR-based markers. Genome, 2002, 45: l150-l156[本文引用:1][JCR: 1.668]
[18]
戴小军, 杨远柱, 周亮, 梁满中, 胡小淳, 陈良碧. 抗稻瘟病水稻资源抗性基因Pi-ta、Pi-b、Pi9以及Pikm的分布研究. 生命科学研究, 2012, 16: 340-344DaiX J, YangY Z, ZhouL, LiangM Z, HuX C, ChenL B. Distribution research of blast resistance genes Pi-ta, Pi-b, Pi-9 and Pikm in blast-resistant rice resources. Life Sci Res, 2012, 16: 340-344 (in Chinese with English abstract)[本文引用:1][CJCR: 0.59]
[19]
LiW, LeiC L, ChengZ J, JiaY L, HuangD Y, WangJ L, WangJ K, ZhangX, SuN, GuoX P, ZhaiH Q, WanJ M. Identification of SSR markers for a broad-spectrum blast resistance gene Pi20(t) for marker-assisted breeding. Mol Breed, 2008, 22: 141-149[本文引用:1][JCR: 3.251]
[20]
王忠华, 贾育林, 吴殿星, 夏英武. 水稻抗稻瘟病基因Pi-ta的分子标记辅助选择. 作物学报, 2004, 30: 1259-1265WangZ H, JiaY L, WuD X, XiaY W. Molecular marker-assisted selection of the rice blast resistance gene Pi-ta. Acta Agron Sin, 2004, 30: 1259-1265 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[21]
FjellstromR, Conaway-BormansC A, McClungA M, MarchettiM A, ShankA R, ParkW D. Development of DNA markers suitable for marker assisted selection of three Pi genes conferring resistance to multiple Pyricularia grisea pathotypes. Crop Sci, 2004, 44: 1790-1798[本文引用:1][JCR: 1.513]
[22]
杨杰, 杨金欢, 王军, 范方军, 朱金燕, 曹卿, 田胜尼, 仲维功. 稻瘟病抗病基因Pi-ta和Pi-b在中国水稻地方品种中的分布. 华北农学报, 2011, 26(3): 1-6YangJ, YangJ H, WangJ, FanF J, ZhuJ Y, CaoQ, TianS N, ZhongW G. Distribution of two blast resistant genes Pita and Pib in land race rice in China. Acta Agric Boreali-Sin, 2011, 26(3): 1-6 (in Chinese with English abstract)[本文引用:2][CJCR: 0.951]
[23]
李进斌, 李成云, 陈艳, 雷财林, 凌忠专. 二十二个抗稻瘟病基因在云南的利用价值评价. 植物保护学报, 2005, 6: 113-119LiJ B, LiC Y, ChenY, LeiC L, LingZ Z. Utilization value of twenty-two blast resistance genes in Yunnan. Acta Phytophy Sin, 2005, 6: 113-119 (in Chinese with English abstract)[本文引用:1]
[24]
王国珍, 茹庆华, 高立原, 李华. 日本水稻抗瘟基因对宁夏稻瘟病菌的抗性研究. 西北农业学报, 1997, 6(3): 1-4WangG Z, RuQ H, GaoL Y, LiH. Studies on genes for resistance blast fungus in Ningxia region. Acta Agric Boreali-Occident Sin, 1997, 6(3): 1-4 (in Chinese with English abstract)[本文引用:1]
[25]
陆凡, 陈志谊, 刘永锋, 郑小波. 江苏省稻瘟病菌毒性的群体结构分析. 植物保护学报, 2002, 29: 289-294LuF, ChenZ Y, LiuY F, ZhengX B. Analysis of population virulences of Magnaporthe grisea in Jiangsu Province. Acta Phytophy Sin, 2002, 29: 289-294 (in Chinese with English abstract)[本文引用:1]
[26]
任金平, 郭晓莉, 郑民, 杨晓辉, 刘晓梅, 李莉, 孙辉. 吉林省稻瘟病菌对水稻品种的致病谱研究. 吉林农业科学, 2006, 31(6): 35-37RenJ P, GuoX L, ZhengM, YangX H, LiuX M, LiL, SunH. Studies on pathogenic spectrum of rice varieties with Magnaporth grisea in Jilin Province. J Jilin Agric Sci, 2006, 31(6): 35-37 (in Chinese with English abstract)[本文引用:1][CJCR: 0.518]
[27]
LiuZ B, GaoQ R, WangR X, QiaoX L, QiuX M. Application of multiplex PCR to studies on plant biology. Mol Plant Breed, 2005, 3: 261-268[本文引用:1][CJCR: 1.23]




{kind=link}
{kind=link}
{kind=link}
{kind=link}