**同等贡献(Contributed equally to this work)收稿日期:2014-04-16 基金:本研究由国家重点基础研究计划(973计划)项目(2014CB138105), 教育部新世纪优秀人才项目(NCET-13-0776), 河南省高校青年骨干教师资助计划项目(2011GGJS-044)和河南省现代农业产业技术体系建设专项(Z2010-01-04)资助;
关键词:硬粒小麦; Lpx-B1位点; 等位变异; 脂肪氧化酶 Molecular Identification of Alleles onLpx-B1 Locus and Lipoxygenase Activity in Durum Wheat (Triticum turgidum L.) ZHANG Fu-Yan1,2,**, SHANG Xiao-Li1,**, WU Pei-Pei1, SONG Shuang1, CHEN Feng1,*, CUI Dang-Qun1,* 1 College of Agronomy / Collaborative Innovation Center of Henan Grain Crops / National Key Laboratory of Wheat & Maize Crop Science, Henan Agricultural University, Zhengzhou 450002, China
2 Henan Key Laboratory of Nuclear Agricultural Sciences / Isotope Institute Co., Ltd, Henan Academy of Sciences, Zhengzhou 450015, China
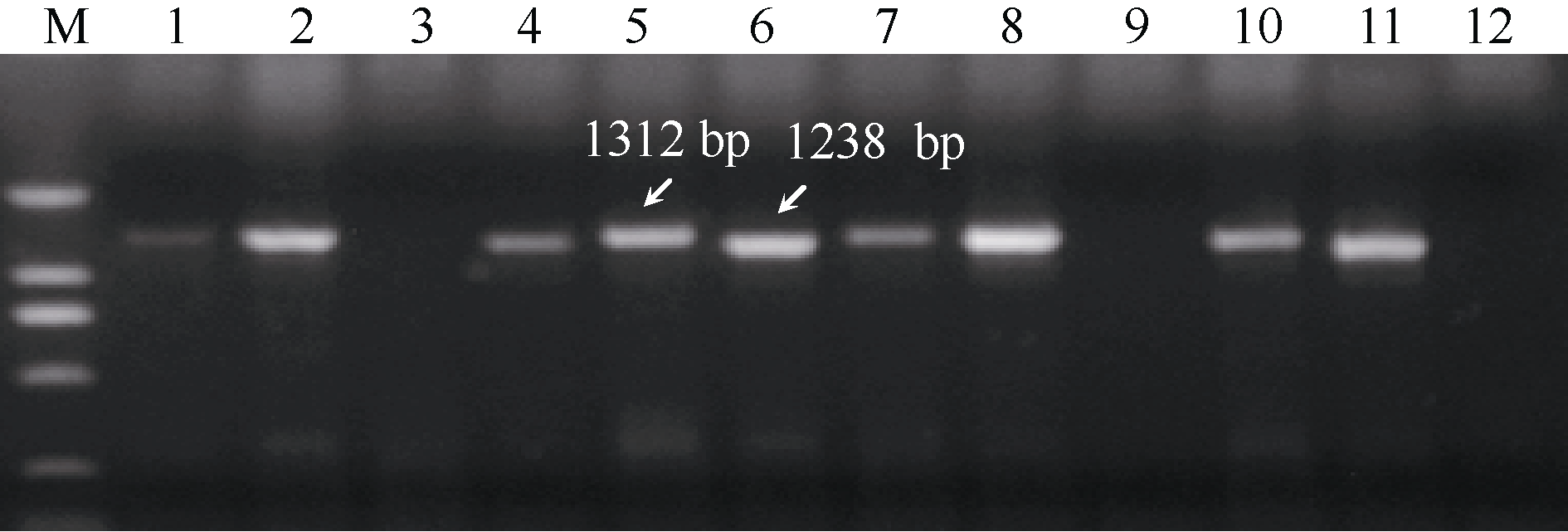
Fund: AbstractLipoxygenase (LOX) has a close relationship to the processing quality of flour products in durum wheat (Triticum turgidum L.). Activity of LOX is greatly influenced by the locus ofLpx-B1. TheLpx-B1 alleles in 167 durum wheat varieties from different countries or regions were identified using gene-specific primers, and theLpx-B1 genotypes were analyzed based on LOX activity data. The LOX activity varied greatly in the 167 varieties, ranging from 0.20 to 7.98 AU min-1 g-1. OnLpx-B1.1 locus, three alleles (Lpx-B1.1a,Lpx-B1.1b, andLpx-B1.1c) were identified with frequencies of 55.1%, 7.8%, and 37.1%, respectively. LociLpx-B1.2andLpx-B1.3were always reciprocally present in varieties, in whichLpx-B1.2 was presented in 146 varieties andLpx-B1.3 in the remaining 21 varieties. This result indicates the allelic possibility ofLpx-B1.2andLpx-B1.3. In the threeLpx-B1.1 genotypes, theLpx-B1.1bvarieties had significantly higher LOX activity than theLpx-B1.1a andLpx-B1.1c varieties, and varieties carryingLpx-B1.1callele possessed the lowest LOX activity. The LOX activity inLpx-B1.3 genotypes was significantly higher than that inLpx-B1.2genotypes. A total of six genotypic combinations were found on the threeLpx-B1 loci in 167 varieties. Among them,Lpx-B1.1b/Lpx-B1.3combination had the highest LOX activity and combinations ofLpx-B1.1c/Lpx-B1.2 andLpx-B1.1c/Lpx-B1.3had the lowest LOX activity. These results are informative to quality breeding of durum wheat.
图3 硬粒小麦中 Lpx-B1位点不同变异类型的LOX活性LOX活性为同类品种的平均值及标准差; 柱形上面的不同字母表示不同变异类型间有显著性差异( P<0.05)。Fig. 3 LOX activities in different Lpx-B1 genotypes of durum wheatLOX activity is the mean of each variety group, and the error bar shows the standard error; Different letters above columns indicate significant difference among different types of variation at P < 0.05.
表4 Table 4 表4(Table 4)
表4 硬粒小麦中 Lpx-B1位点不同基因型组合LOX活性比较 Table 4 Comparison of LOX activities of durum wheat varieties with different genotype combinations
基因型组合 Genotype combination
LOX类型 LOX type
品种数 No. of varieties
平均LOX活性 Mean LOX activity (AU min-1 g-1)
频率 Frequency (%)
Lpx-B1.1b/Lpx-B1.3
H
11
6.03 a
6.6
Lpx-B1.1a/Lpx-B1.3
MH
9
4.27 b
5.4
Lpx-B1.1b/Lpx-B1.2
MH
2
4.17 b
1.2
Lpx-B1.1a/Lpx-B1.2
ML
83
2.30 c
49.7
Lpx-B1.1c/Lpx-B1.3
L
1
1.02 d
0.6
Lpx-B1.1c/Lpx-B1.2
L
61
0.92 d
36.5
H: 高LOX型; MH: LOX偏高型; ML: LOX偏低型; L: 低LOX型。LOX活性后不同字母表示基因型组合类群间有显著差异( P<0.05)。 H: high LOX activity; MH: moderately high LOX activity; ML: moderately low LOX activity; L: low LOX activity. Different letters after the mean LOX activity indicate significant difference among genotype combinations at P < 0.05.
表4 硬粒小麦中 Lpx-B1位点不同基因型组合LOX活性比较 Table 4 Comparison of LOX activities of durum wheat varieties with different genotype combinations
LoiseauJ, VuB L, MacherelM H, DeunffY L. Seed lipoxygeanses: occurrence and functions. Seed Sci Res, 2001, 11: 199-211[本文引用:1][JCR: 1.931]
[2]
汪仁, 沈文飚, 翟虎渠, 万建民. 植物种子脂氧合酶. 植物生理学通讯, 2005, 41: 388-395WangR, ShenW B, ZhaiH Q, WanJ M. Plant seed lipoxygenase. Plant Physiol Commun, 2005, 41: 388-395 (in Chinese with English abstract)[本文引用:2]
[3]
HesslerT G, ThomsonM J, BenscherD, NachitM M, SorrellsM E. Association of a lipoxygenase locus, Lpx-B1, with variation in lipoxygenase activity in durum seeds. Crop Sci, 2002, 42: 1695-1700[本文引用:3][JCR: 1.513]
[4]
ShiibaK, NengishiY, OkadaK, NagaoS. Purification and characterization of lipoxygenase isozymes from wheat germ. Cereal Chem, 1991, 68: 115-122[本文引用:1][JCR: 1.251]
[5]
PastoreD, TronoD, PadalinoL, SimoneS, ValentiD, FonzoN D, PassarellaS. Inhibition by α-tocopherol and L-aseorbate of linoleate hydroperoxidation and β-carotene bleaching activities in durum wheat semolina. J Cereal Sci, 2000, 31: 41-54[本文引用:1][JCR: 2.088]
[6]
郑文寅, 姚大年, 张文明. 脂肪氧化酶及其在小麦品质改良中的研究与应用. 粮食与饲料工业, 2009, (8): 5-6ZhengW Y, YaoD N, ZhangW M. Research and application of lipoxygenase for improvement of wheat quality. Cereal Feed Ind, 2009, (8): 5-6 (in Chinese with English abstract)[本文引用:3]
[7]
BorrelliG M, DeleonardisA M, PlataniC, TroccoliA. Distribution along durum wheat kernel of the components involved in semolina color. J Cereal Sci, 2008, 48: 494-502[本文引用:1][JCR: 2.088]
[8]
王慧, 郑文寅, 樊宏, 汪帆, 王青, 王冠球, 张文明, 姚大年. 不同小麦品种籽粒中LOX活性及基因型和环境互作分析. 中国粮油学报, 2011, 26(1): 11-14WangH, ZhengW Y, FanH, WangF, WangQ, WangG Q, ZhangW M, YaoD N. Lipoxygenase activity and its genotype and environment interactions for different wheat varietier. J Chin Cereals Oils Assoc, 2011, 26(1): 11-14 (in Chinese with English abstract)[本文引用:1]
[9]
LeenhardtF, LyanaB, RockaE, BoussardA, PotusJ, ChanliaudE, RemesyC. Genetic variability of carotenoid concentration, and lipoxygenase and peroxidase activities among cultivated wheat species and bread wheat varieties. Eur J Agron, 2006, 25: 170-176[本文引用:1][JCR: 2.8]
[10]
HartG E, LangstonP J. Chromosome location and evolution of isozyme structural genes in hexaploid wheat. Heredity, 1977, 39: 263-277[本文引用:1][JCR: 4.11]
[11]
HsiehC C, McDonaldCE. Isolation of lipoxygenase isoenzymes from ?our of durum wheat endosperm. Cereal Chem, 1984, 61: 392-398[本文引用:1][JCR: 1.251]
[12]
GarbusI, CarreraA D, DubcovskyJ, EcheniqueV. Physical mapping of durum wheat lipoxygenase genes. J Cereal Sci, 2009, 50: 67-73[本文引用:2][JCR: 2.088]
[13]
PsheniehnikovaT A, OsipovaS V, PermyakovaM D, MitrofanovaT N, TrufanovV A, LohwasserU, RöderM, BörnerA. Mapping of quantitative trait loci (QTL) associated with activity of disulfide reductase and lipoxygenase in grain of bread wheat. Russ J Genet, 2008, 44: 567-574[本文引用:2][JCR: 0.427]
[14]
CarreraA, EcheniqueV, ZhangW, HelgueraM, MantheyF, SchragerA, PiccaA, CervigniG, DubcovskyJ. A deletion at the Lpx-B1 locus is associated with low lipoxygenase activity and improves pasta color in durum wheat (Triticum turgidum ssp. durum). J Cereal Sci, 2007, 45: 67-77[本文引用:2][JCR: 2.088]
[15]
VerlottaA, SimoneV D, MastrangeloA M, CattivelliL, PapaR, TronoD. In sight into durum wheat Lpx-B1: a small gene family coding for the lipoxygenase responsible for carotenoid bleaching in mature grains. BMC Plant Biol, 2010, 10: 263[本文引用:6][JCR: 4.354]
[16]
FengB, DongZ Y, XuZ B, WangD W, WangT. Molecular characterization of a novel type of lipoxygenase (LOX) gene from common wheat (Triticum aestivum L. ). Mol Breed, 2012, 30: 113-124[本文引用:2][JCR: 3.251]
[17]
GengH W, XiaX C, ZhangL P, QuY Y, HeZ H. Development of functional markers for a lipoxygenase gene TaLox-B1 on chromosome 4BS in common wheat. Crop Sci, 2012, 52: 568-576[本文引用:1][JCR: 1.513]
[18]
GarbusI, SoresiD, RomeroJ, EcheniqueV. Identification, mapping and evolutionary course of wheat lipoxygenase-1 genes located on the A genome. J Cereal Sci, 2013, 58: 298-304[本文引用:2][JCR: 2.088]
[19]
FengB, DongZ Y, XuZ B, AnX L, QinH J, WuN, WangD W, WangT. Molecular analysis of lipoxygenase (LOX) genes in common wheat and phylogenetic investigation of LOX proteins from model and crop plants. J Cereal Sci, 2010, 52: 387-394[本文引用:2][JCR: 2.088]
[20]
LeenhardtF, LyanB, RockE, BoussardA, PotusJ, ChanliaudE, RemesyC. Wheat lipoxygenase activity induces greater loss of carotenoids than vitamin E during bread making. J Agric Food Chem, 2006, 54: 1710-1715[本文引用:1][JCR: 2.906]
[21]
BorrelliG M, Ficco D B M, FonzoN D, FaresC. Effects of lipoxygenase and of chemical oxidising agent potassium iodate on rheological properties of durum dough. Int J Food Sci Technol, 2006, 41: 639-645[本文引用:1][JCR: 1.24]
[22]
郑文寅, 汪帆, 司红起, 张文明, 姚大年. 普通小麦籽粒LOX、PPO活性和类胡萝卜素含量变异及对全麦粉色泽的影响. 中国农业科学, 2013, 46: 1087-1094ZhengW Y, WangF, SiH Q, ZhangW M, YaoD N. Variations of LOX and PPO activities and carotenoid content as well as their influence on whole flour color in common wheat. Sci Agric Sin, 2013, 46: 1087-1094 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[23]
ChenF, HeZ H, XiaX C, LillemoM, MorrisC. A new puroindoline b mutation presented in Chinese winter wheat cultivar Jingdong 11. J Cereal Sci, 2005, 42: 267-269[本文引用:1][JCR: 2.088]
[24]
BorrelliG M, TroeeoliA, FonzoN D, FaresC. Durum wheat lipoxygenase activity and other quality parameters that affect pasta color. Cereal Chem, 1999, 76: 335-340[本文引用:1][JCR: 1.251]
[25]
赵丹, 张玉荣, 林家永, 周显青. 小麦储藏品质评价指标研究进展. 粮食与饲料工业, 2012, (2): 10-14ZhaoD, ZhangY R, LinJ Y, ZhouX Q. Research on quality evaluation of wheat storage. Cereal Feed Ind, 2012, (2): 10-14 (in Chinese with English abstract)[本文引用:1]
[26]
TrufanovV A, PermyakovaM D, PshenichnikovaT A, ErmakovaM F, DavydovV A, PermyakovA V, BerezovskayaE V. The effect of intercultivar substitution of wheat Triticum aestivum L. chromosomes on lipoxygenase activity and its correlation with the technological properties of flour. Appl Biochem Microbiol, 2007, 43: 91-97[本文引用:1][JCR: 0.689]
[27]
PermyakovaM D, TrufanovV A, PshenichnikovaT A, ErmakovaM F. Role of lipoxygenase in the determination of wheat grain quality. Appl Biochem Microbiol, 2010, 46: 87-92[本文引用:1]
[28]
ŽilićS, DodigD, Šukalović V H T, MaksimovićM, SaratlićG, ŠkrbićB. Bread and durum wheat compared for antioxidants contents, and lipoxygenase and peroxidase activities. Int J Food Sci Tech, 2010, 45: 1360-1367[本文引用:1][JCR: 1.24]
[29]
GengH W, ZhangY, HeZ H, ZhangL P, AppelsR, QuY Y, XiaX C. Molecular markers for tracking variation in lipoxygenase activity in wheat breeding. Mol Breed, 2011, 28: 117-126[本文引用:1][JCR: 3.251]
LiuY N, HeZ H, AppelsR, XiaX C. Functional markers in wheat: current status and future prospects. Theor Appl Genet, 2012, 125: 1-10[本文引用:2][JCR: 3.658]
 , 崔党群
, 崔党群





{kind=link}
{kind=link}
{kind=link}