关键词:小麦-黑麦种质; 农艺性状; GISH; FISH; 小黑麦; 遗传多样性 Main Agronomic Traits of 390 Wheat-Rye Derivatives and GISH/FISH Identification of Their Outstanding Materials LUO Qiao-Ling1,4, ZHENG Qi2, XU Yun-Feng1, LI Li-Hui3, HAN Fang-Pu2, XU Hong-Xing1, LI Bin2, MA Peng-Tao1, AN Diao-Guo1,* 1 Center for Agricultural Resources Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Shijiazhuang, Hebei 050022, China
2 State Key Laboratory of Plant Cell and Chromosome Engineering, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China
3 Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China
4 University of Chinese Academy of Sciences, Beijing 100049, China
Fund: AbstractTransferring desirable genes of rye (Secale cereale L.) into common wheat (Triticum aestivum L.) can widen the genetic basis and enrich genetic variation of wheat. We analyzed 390 wheat-rye derivatives in this study. Large ranges of variation were found according to six main agronomic traits, indicating an abundant genetic diversity in these germplasms. Compared to ten major wheat cultivars, more than 90% of the wheat-rye derivatives were superior in spike length and tiller number, more than 60% were superior in spikelet number, and about 30% had higher kernel number per spike and thousand-grain weight. Eight representative materials with desirable agronomic traits were tested with genomicin situhybridization (GISH) and multicolor fluorescentin situhybridization (mc-FISH). The result showed that among eight outstanding materials three were hexaploid triticales (AABBRR) and two were octoploid triticales (AABBDDRR); another one was the 1RS·1BL translocation line; and the remaining two lines contained neither chromosome nor chromosome fragments of rye. Interestingly, the rye chromosomes were not completely the same between the hexaploid triticale and the octoploid triticale. A pair of rye chromosomes in the octoploid triticale was smaller than usual, whereas the hexaploid triticale had no such small chromosomes. We also found that different triticale materials had different GISH banding patterns of chromosomes 4R. These results provide a basis for application of the wheat-rye derivatives in wheat breeding.
Keyword:Wheat-rye derivatives; Agronomic trait; GISH; FISH; Triticale; Genetic diversity Show Figures Show Figures
2 结果与分析2.1 390份小麦-黑麦种质材料主要农艺性状的分析390份小麦-黑麦种质的分蘖数、穗长、小穗数、穗粒数和千粒重变异范围很大, 分蘖数相差27.8, 穗长相差9.0 cm, 小穗数相差19.2, 穗粒数相差74.0粒, 千粒重相差39.5 g (表1)。大部分材料的性状值集中在 表1 Table 1 表1(Table 1)
表1 390份小麦-黑麦种质材料与10份小麦主栽品种主要农艺性状比较 Table 1 Comparison of 390 wheat-rye derivatives with 10 major wheat cultivars in main agronomic traits
性状 Trait
主栽品种 Major cultivars
小麦-黑麦种质 Wheat-rye derivatives
优于主栽品种的小麦-黑麦种质数1) No. of wheat-rye derivatives superior to major cultivars1)
平均值 Mean
范围 Range
平均值 Mean
范围 Range
优于平均值 vs. mean
优于最大值 vs. maximum
优于最小值 vs. minimum
分蘖数 TN
9.3
7.0-10.7
14.3*
5.0-32.8
355 (91.0%)
317 (81.3%)
385 (98.7%)
穗长SL (cm)
8.6
7.2-10.2
12.5*
8.3-17.3
379 (97.2%)
348 (89.2%)
390 (100.0%)
小穗数SN
18.9
18.3-20.5
22.0*
15.8-35.0
254 (65.1%)
224 (57.4%)
357 (91.5%)
不育小穗数 SSN
0.4
0-1.0
0.8*
0-8.3
104 (26.7%)
260 (66.7%)
—
穗粒数KNS
54.9
52.4-65.0
54.1*
17.8-91.8
99 (25.4%)
43 (11.0%)
204 (52.3%)
千粒重TKW (g)
38.0
32.9-43.3
36.6*
21.0-60.5
137 (35.1%)
38 (9.7%)
293 (75.1%)
10个小麦主栽品种分别是济麦22、石新828、科农199、良星99、石4185、石麦15、冀5265、山农21、烟农19和邯7086。*表示小麦-黑麦种质材料显著优于主栽品种 ( P<0.05)。1)括号中数据为超过主栽品种的百分数。优于主栽品种平均值的不育小穗数指不育小穗数为0的材料数和比例。 The 10 major wheat cultivars were Jimai 22, Shixin 828, Kenong 199, Liangxing 99, Shi 4185, Shimai 15, Ji 5265, Shannong 21, Yannong 19, and Han 7086. TN: tiller number; SL: spike length; SN: spikelet number; SSN: sterile spikelet number; KNS: kernel number per spike; TKW: 1000-kernel weight.* indicates significantly superior to the main cultivars at P < 0.05.1)Data in parentheses are the percentages. SSN superior to the mean of major cultivars indicates that the number and percentage of wheat-rye derivatives with zero sterile spikelet.
表1 390份小麦-黑麦种质材料与10份小麦主栽品种主要农艺性状比较 Table 1 Comparison of 390 wheat-rye derivatives with 10 major wheat cultivars in main agronomic traits
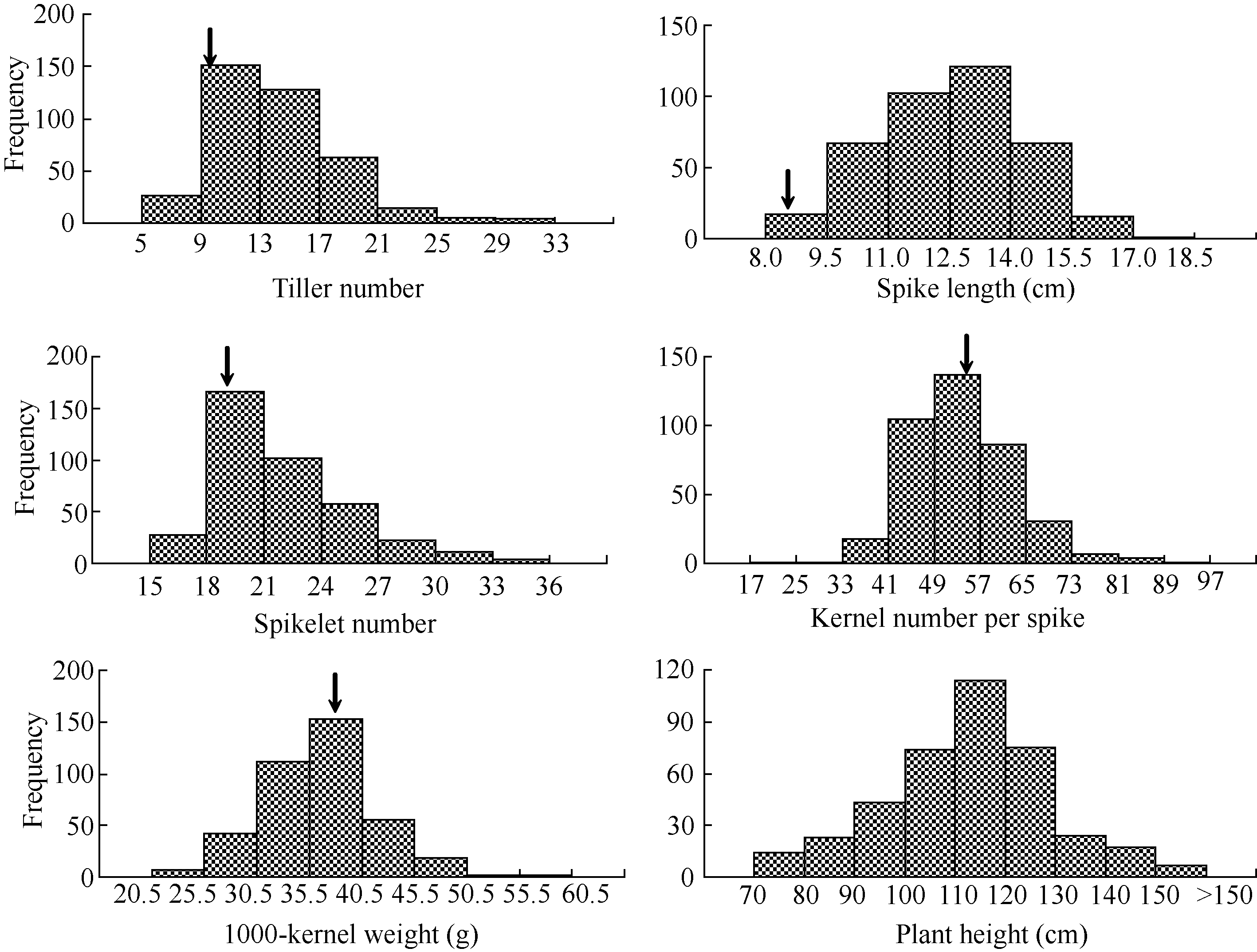
一定范围内, 如87.7%的材料分蘖数在9~21区间, 91.5%的材料穗长在9.5~15.5 cm区间, 83.3%的材料小穗数为18~27, 84.1%的材料穗粒数为41~65, 82.1%的材料千粒重集中在30.5~45.5 g范围内。此外, 390份材料株高变异范围为71.0~172.6 cm, 极差为101.6 cm, 其中67.4%的材料为100~130 cm, 还有7份材料株高超过150 cm, 仅14份材料(3.6%)的株高为70~80 cm (图1)。 超过90%的小麦-黑麦种质材料表现为分蘖数和穗长显著优于小麦主栽品种, 并且385份材料(98.7%)的分蘖数超过主栽品种的最小值7.0 (科农199); 所有材料的穗长都大于主栽品种的最小值7.2 cm (科农199); 小穗数也有明显优势, 其中254份材料(65.1%)显著优于主栽品种, 357份材料(91.5%)的小穗数高于主栽品种的最小值18.3 (山农21)。良星99、山农21和烟农19的不育小穗数都为0, 石新828、石麦15、冀5265和科农199都为1, 而260份(66.7%)小麦-黑麦种质材料的不育小穗数小于1 (表1)。穗粒数和千粒重两性状, 小麦-黑麦种质材料的优势相对较弱, 但也有43份材料(11.0%)的穗粒数大于主栽品种最大值65.0 (山农21), 204份材料(52.3%)大于主栽品种最小值52.4 (石新828); 38份材料(9.7%)的千粒重大于主栽品种的最大值43.3 g (良星99), 293份材料(75.1%)大于主栽品种最小值32.9 g (石4185)。 图1 Fig. 1
图1 390份小麦-黑麦种质6个农艺性状的频次分布箭头示10份主栽品种的平均值。Fig. 1 Distributions of frequency in six agronomic traits of 390 wheat-rye derivativesThe arrows show the means of 10 major wheat cultivars.
表2 Table 2 表2(Table 2)
表2 8份代表性材料的主要农艺性状 Table 2 Main agronomic traits of eight representative wheat-rye derivatives
材料 Line
分蘖数 TN
穗长 SL (cm)
小穗数 SN
穗粒数 KNS
千粒重 TKW (g)
株高 PH (cm)
产量 Yield (kg hm-2)
R1162
17.0
16.2
34.4
51.4
43.2
155.0
8684
R2203
14.8
15.9
26.6
46.2
45.0
134.0
7253
R2221
19.4
16.0
27.8
65.3
43.2
140.8
7645
R1017
16.6
11.4
17.8
35.8
60.5
127.0
5196
R1103
13.4
15.5
35.0
47.0
55.0
149.0
4683
R1147
32.8
11.9
19.8
49.0
32.9
105.2
3437
R2209
11.6
14.8
29.2
91.8
40.3
152.0
5959
R1156
19.0
9.6
19.0
54.8
37.2
87.0
4032
TN: tiller number; SL: spike length; SN: spikelet number; KNS: kernel number per spike; TKW: 1000-kernel weight; PH: plant height.
表2 8份代表性材料的主要农艺性状 Table 2 Main agronomic traits of eight representative wheat-rye derivatives
图2 8份小麦-黑麦种质材料的GISH和mc-FISH鉴定A~F依次为小麦-黑麦种质材料R1103、R1162、R2203、R2221、R1147和R1156。A1~F1是以黑麦基因组DNA为探针(绿色), 中国春基因组DNA为封阻的GISH鉴定结果, 其中B1和D1中的十字星号示无端带的2条黑麦染色体, C1和D1中的箭头示1对黑麦的小染色体。 A2~F2是以重复序列p As1 (红色)和p Sc119.2 (绿色)为探针的mc-FISH鉴定结果, C2和D2中的问号示不能确定归属的染色体。Fig. 2 Identification of GISH and mc-FISH of eight wheat-rye derivativesA-F are wheat-rye derivatives R1103, R1162, R2203, R2221, R1147, and R1156, respectively. A1-F1 are GISH results with rye genomic DNA as a probe (green) and Chinese Spring DNA as a blocker. The cross stars in B1 and D1 show the two rye chromosomes without GISH binding pattern, and the arrows in C1 and D1 show a pair of small rye chromosomes. A2-F2 are mc-FISH results on the same metaphase after GISH analysis by p As1 (red) and p Sc119.2 (green) simultaneously. The question marks in C2 and D2 show the undetermined chromosomes.
郝晨阳, 王兰芬, 张学勇, 游光霞, 董玉琛, 贾继增, 刘旭, 尚勋武, 刘三才, 曹永生. 我国育成小麦品种的遗传多样性演变. 中国科学C辑·生命科学, 2005, 35: 408-415HaoC Y, WangL F, ZhangX Y, YouG X, DongY C, JiaJ Z, LiuX, ShangX W, LiuS C, CaoY S. Changes of genetic diversity of wheat varieties released in China. Sci China Ser C: Life Sci, 2005, 35: 408-415 (in Chinese)[本文引用:1]
[2]
HarperJ, ArmsteadI, ThomasA, JamesC, GasiorD, BisagaM, RobertsL, KingI, KingJ. Alien introgression in the grasses Lolium perenne (perennial ryegrass) and Festuca pratensis (meadow fescue): the development of seven monosomic substitution lines and their molecular and cytological characterization. Ann Bot-London, 2011, 107: 1313-1321[本文引用:1][JCR: 3.449]
[3]
安调过, 许红星, 许云峰. 小麦远缘杂交种质资源创新. 中国生态农业学报, 2011, 19: 1011-1019AnD G, XuH X, XuY F. Enhancement of wheat distant hybridization germplasm. Chin J Eco-Agric, 2011, 19: 1011-1019 (in Chinese with English abstract)[本文引用:2][CJCR: 0.795]
[4]
CarverB F, RayburnA L. Comparison of related wheat stocks possessing 1B or 1RS·1BL chromosomes agronomic performance. Crop Sci, 1994, 34: 1505-1510[本文引用:1][JCR: 1.513]
[5]
WangC M, ZhengQ, LiL H, NiuY C, WangH B, LiB, ZhangX T, XuY F, AnD G. Molecular cytogenetic characterization of a new T2BL center dot 1RS wheat-rye chromosome translocation line resistant to stripe rust and powdery mildew. Plant Dis, 2009, 93: 124-129[本文引用:1][JCR: 2.455]
[6]
ZhouJ P, ZhangH Y, YangZ J, LiG R, HuL J, LeiM P, LiuC, ZhangY, RenZ L. Characterization of a new T2DS·2DL-?R translocation triticale ZH-1 with multiple resistances to diseases. Gen Resour Crop Evol, 2012, 59: 1161-1168[本文引用:1]
[7]
AnD G, ZhengQ, ZhouY L, MaP T, LvZ L, LiL H, LiB, LuoQ L, XuH X, XuY F. Molecular cytogenetic characterization of a new wheat-rye 4R chromosome translocation line resistant to powdery mildew. Chrom Res, 2013, 21: 419-432[本文引用:1]
[8]
孙元枢. 中国小黑麦遗传育种研究与应用. 杭州: 浙江科学技术出版社, 2002. pp14-24SunY S. Triticale Genetic Breeding and Utilization in China. Hangzhou: Zhejiang Science and Technology Press, 2002. pp14-34(in Chinese)[本文引用:2]
[9]
SebestaE E, WoodE A, PorterD R, WebsterJ A, SmithE L. Registration of amigo wheat germplasm resistant to greenbug. Crop Sci, 1995, 35: 293-293[本文引用:2][JCR: 1.513]
[10]
LukaszewskiA J. Frequency of 1RS. 1AL and 1RS. 1BL translocations in United-States wheats. Crop Sci, 1990, 30: 1151-1153[本文引用:1][JCR: 1.513]
[11]
RabinovichS V. Importance of wheat-rye translocations for breeding modern cultivars of Triticum aestivum L. Euphytica, 1998, 100: 323-340[本文引用:2][JCR: 1.643]
[12]
SchneiderA, Molnar-LangM. Detection of the 1RS chromosome arm in Martonvásár wheat genotypes containing 1BL·1RS or 1AL·1RS translocations using SSR and STS markers. Acta Agron Hung, 2009, 57: 409-416[本文引用:1]
[13]
尚海英, 郑有良, 魏育明, 吴卫. 黑麦属基因资源研究进展. 麦类作物学报, 2003, 23(1): 86-89ShangH Y, ZhengY L, WeiY M, WuW. Advantage of study on the genetic resources of Secale. J Triticeae Crops, 2003, 23(1): 86-89 (in Chinese with English abstract)[本文引用:1]
[14]
郑琪. 蓝粒小麦易位系的分子细胞遗传学分析及应用. 中国科学院研究生院博士学位论文, 北京, 2006. pp27-28ZhengQ. Molecular Cytogenetic Verification and Utilization of Blue-Grained Wheat Translocation Lines. PhD Dissertation of Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China, 2006. pp27-28 (in Chinese with English abstract)[本文引用:1]
[15]
LeH T, ArmstrongK C, MikiB. Detection of rye DNA in wheat-rye hybrids and wheat translocation stocks using total genomic DNA as a probe. Plant Mol Biol Rep, 1989, 7: 150-158[本文引用:1][JCR: 5.319]
[16]
CaiX, JonesS S, MurrayT D. Molecular cytogenetic characterization of Thinopyrum genomes conferring perennial growth habit in wheat-Thinopyrum amphiploids. Plant Breed, 2001, 120: 21-26[本文引用:1][JCR: 1.175]
[17]
MukaiY, NakaharaY, YamamotoM. Simultaneous discrimination of three genomes in hexaploid wheat by multicolor fluorescence in situ hybridization using total genomic and highly repeated DNA probes. Genome, 1993, 36: 489-494[本文引用:2][JCR: 1.668]
[18]
PedersenC, LangridgeP. Identification of entire chromosome complement of bread wheat by two-colour FISH. Genome, 1997, 40: 589-593[本文引用:1][JCR: 1.668]
[19]
NagyE D, Molnár-LángM, LincG, LángL. Identification of wheat-barley translocations by sequential GISH and two-colour FISH in combination with the use of genetically mapped barley SSR markers. Genome, 2002, 45: 1238-1247[本文引用:1][JCR: 1.668]
[20]
李立会, 李秀全. 小麦种质资源描述规范和数据标准. 北京: 中国农业出版社, 2006LiL H, LiX Q. Stand ard of Description and Data in Wheat Germplasm Resources. Beijing: China Agriculture Press, 2006 (in Chinese)[本文引用:1]
[21]
HanF P, GaoZ, YuW C, BirchlerJ A. Minichromosome analysis of chromosome pairing, disjunction, and sister chromatid cohesion in maize. Plant Cell, 2007, 19: 3853-3863[本文引用:1][JCR: 9.251]
[22]
RayburnA L, GillB S. Isolation of D-genome specific repeated DNA sequence from Aegilops squarrosa. Plant Mol Biol Rep, 1987, 4: 102-109[本文引用:1][JCR: 5.319]
[23]
McIntyreC L, PereiraS, MoranL B. New Secale cereale (rye) DNA deribatives for detection of rye chromosome segments in wheat. Genome, 1990, 33: 635-640[本文引用:1][JCR: 1.668]
[24]
CuadradoA, SchwarzacherT, The chromosomal organization of simple sequence repeats in wheat rye genomes. Chromosome, 1998, 107: 587-594[本文引用:1][JCR: 2.847]
[25]
田纪春, 邓志英, 胡瑞波, 王延训. 不同类型超级小麦产量构成因素及籽粒产量的通径分析. 作物学报, 2006, 32: 1699-1705TianJ C, DengZ Y, HuR B, WangY X. Yield components of super wheat cultivar with different types and the path coefficient analysis on grain yield. Acta Agron Sin, 2006, 32: 1699-1705 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[26]
何中虎, 夏先春, 陈新民, 庄巧生. 中国小麦育种进展与展望. 作物学报, 2011, 37: 202-215HeZ H, XiaX C, ChenX M, ZhuangQ S. Progress of wheat breeding in China and the future perspective. Acta Agron Sin, 2011, 37: 202-215 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[27]
符书兰. 普通小麦(Triticum aestivum L. )与黑麦(Secale cereale L. )异源多倍体及异染色体系的分子细胞学研究. 四川农业大学博士学位论文, 四川雅安, 2009. pp31-34FuS L. Molecular and Cytobiology Study on Allopolyploid and Alien Chromosome Lines of Triticum aestivum L. and Secale cereale L. PhD Dissertation of Sichuan Agricultural University, Ya’an, China, 2009. pp31-34 (in Chinese with English abstract)[本文引用:1]
[28]
BentoM, GustafsonJ P, ViegasW, SilvaM. Size matters in Triticeae polyploids: larger genomes have higher remodeling. Genome, 2011, 54: 175-183[本文引用:1][JCR: 1.668]
[29]
任正隆, LelleyT, RöbbelenG. 小麦和黑麦染色体在小黑麦与小麦杂种的不同世代群体中的传递. 遗传学报, 1991, 18: 161-167RenZ L, LelleyT, RöbbelenG. Transmission of the wheat and rye chromosomes in octoploid triticale × common wheat populations. Acta Genet Sin, 1991, 18: 161-167 (in Chinese with English abstract)[本文引用:2]
[30]
符书兰, 唐宗祥, 任正隆, 小麦-黑麦附加系的创制及5R抗白粉病新基因的发现. 遗传, 2011, 33: 1258-1262FuS L, TangZ X, RenZ L. Establishment of wheat-rye addition lines and De Novo powdery mildew resistance gene from chromosome 5R. Hereditas (Beijing), 2011, 33: 1258-1262 (in Chinese with English abstract)[本文引用:1]
[31]
李方安, 唐宗祥, 符书兰. 新型小麦-黑麦6R附加系的创制及其白粉病抗性基因向小麦中的渗进. 麦类作物学报, 2013, 34: 33-37LiF A, TangZ X, FuS L. Development and identification of the novel wheat-rye 6R addition line with resistant gene to powdery mildew. J Triticeae Crops, 2013, 34: 33-37 (in Chinese with English abstract)[本文引用:1]
[32]
姚景侠, 钟少斌, 张德玉. 六倍体小黑麦抗白粉病基因导入普通小麦的研究. 见: 庄巧生, 杜振华主编. 中国小麦育种研究进展(1991-1995). 北京: 中国农业出版社, 1996. pp331-338YaoJ X, ZhongS B, ZhangD Y. Research of transferring resistant genes to , 1996. pp331-338(in Chinese)[本文引用:1]
[33]
SaulescuN N, IttuG, CiucaM, IttuM, SerbanG, MustateaP. Transferring useful rye genes to wheat, using triticale as a bridge. Czech J Genet Plant, 2011, 47: S56-S62[本文引用:1][JCR: 0.385]
[34]
KimW, JohnsonJ W, BaenzigerP S, LukaszewskiA J, GainesC S. Agronomic effect of wheat-rye translocation carrying rye chromatin (1R) from different sources. Crop Sci, 2004, 44: 1254-1258[本文引用:2][JCR: 1.513]
[35]
GrayboschR A. Uneasy unions: quality effects of rye chromatin transfers to wheat. J Cereal Sci, 2001, 33: 3-16[本文引用:1][JCR: 2.088]
[36]
KumlayA M, BaenzigerP S, GillK S, SheltonD R, GrayboschR A, LukaszewskiA J, WesenbergD M. Understand ing the effect of rye chromatin in bread wheat. Crop Sci, 2003, 43: 1643-1651[本文引用:1][JCR: 1.513]
[37]
周建平, 张怀渝, 张怀琼, 任正隆. 小麦新品种川农12号中外源染色质的分子细胞遗传学检测. 麦类作物学报, 2006, 26(4): 20-22ZhouJ P, ZhangH Y, ZhangH Q, RenZ L. Molecular cytogenetics identification of alien chromatin in Chuannong 12. J Triticeae Crops, 2006, 26(4): 20-22 (in Chinese with English abstract)[本文引用:1]
[38]
任天恒, 晏本菊, 张怀琼, 任正隆. 1RS·1BL易位染色体对小麦开花后叶片延绿特性的影响. 麦类作物学报, 2009, 29: 419-423RenT H, YanB J, ZhangH Q, RenZ L. Effect of 1RS·1BL translocation chromosome on stay green trait in common wheat (Triticum aestivum L. ). J Triticeae Crops, 2009, 29: 419-423 (in Chinese with English abstract)[本文引用:1]
[39]
樊小莉, 王静, 张玮, 纪军, 张相岐, 李俊明. 小麦高产抗病新种质9204R的分子细胞学鉴定. 麦类作物学报, 2011, 31: 611-616FanX L, WangJ, ZhangW, JiJ, ZhangX Q, LiJ M. Identification of wheat-rye translocation line 9204R with desirable agronomic traits and stripe rust resistance. J Triticeae Crops, 2011, 31: 611-616 (in Chinese with English abstract)[本文引用:1]
[40]
任正隆. 遗传转移的MADI过程. 四川农业大学学报, 1990, 8(1): 1-6RenZ L. The MADI process for genetic transfer. J Sichuan Agric Univ, 1990, 8(1): 1-6 (in Chinese with English abstract)[本文引用:1][CJCR: 0.5]




{kind=link}
{kind=link}